
Проблемы Эволюции |
2003 г. Палеонтологический журнал. В печати.
Фотоальбом "Ископаемые прокариоты"
Исследования ископаемых бактерий приводит к заключению, что характер седиментации в эпиконтинентальных бассейнах существенно зависит от обилия микроорганизмов в их водной толще. По состоянию знаний на сегодня выясняется гораздо более раннее, чем обычно считается, появление эвкариот, целомат и т.д., и гораздо более ранняя оксигенезация атмосферы.
Ключевые слова: fossil bacteria, evolution of biosphere, sedimentology.
В настоящей статье, отражающей содержание доклада в МГУ (2002 г.), мне хотелось бы кратко остановиться на двух важных проблемах: влиянии микроорганизмов на седиментогенез в эпиконтинентальных бассейнах прошлого и палеонтологических доказательствах появления тех или иных групп органического мира в докембрийской истории Земли.
Последние 15-20 лет можно считать временем, весьма значимым для начала осознания многих эволюционных, седиментологических и биосферных вопросов в связи с развитием геомикробиологии, т.е. науки о взаимодействии микроорганизмов с различными материалами, включая горные породы (Banfiеld, Nealson, 1997), интенсивным изучением цианобактериальных матов (Заварзин, 1994) и более простых биопленок и, наконец, становлением бактериальной палеонтологии (Розанов, Заварзин, 1997). Именно в процессе бактериально-палеонтологических исследований стало ясно, что микроорганизмы, и особенно бактерии, очень часто прекрасно фоссилизируются и великолепно сохраняются в ископаемом состоянии. Начало было положено Т.С. Баргхурном, его коллегами и учениками (Barghoorn, Tylor, 1965; Sсhopf, 1983), но долгое время считалось, что перкрасная сохранность характерна для микроорганизмов только в кремнистых породах.
Электронная микроскопия для исследований ископаемых микроорганизмов, как ни парадоксально, почти не применялась, поскольку знаменитые «кремнистые биоты» и акритархи докембрия традиционно могли в основном изучаться в шлифах и мацерациях с помощью оптического микроскопа.
Широкое использование сканирующего микроскопа фактически началось с изучения микроорганизмов в древних фосфоритах Хубсугула (нижний кембрий, томмотский ярус, Монголия) (Розанов, Жегалло, 1989; Zhegallo et al., 2000). Опыт предыдущей работы с фосфатными окаменелостями (так называемыми мелкими скелетными ископаемыми) из пограничных отложений докембрия и кембрия (Розанов и др., 1969) навел на мысль о необходимости предварительного непродолжительного (обычно несколько секунд или минут) травления породы слабыми кислотами. Именно поэтому в результате резкой разницы в растворимости карбонатов, фосфатов и кремнистой составляющей образуется рельеф, позволяющий прекрасно рассматривать ископаемые объекты (рис. 1).
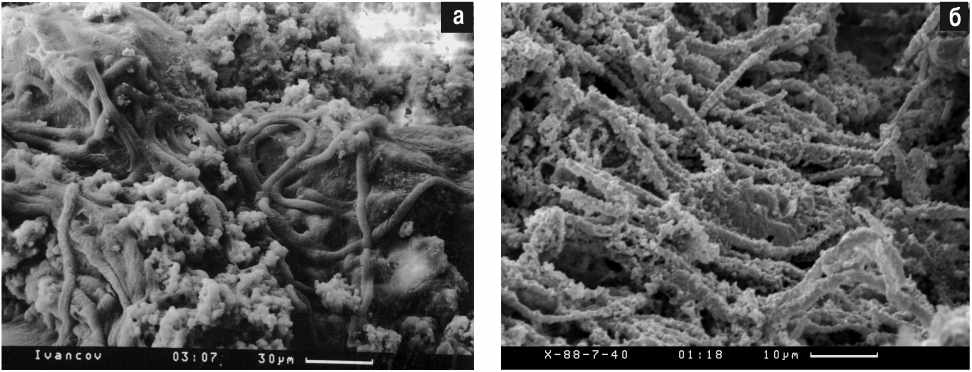
Рис. 1. Современные (а) и раннекембрийские (б) цианобактерии.
Сохранность цианобактерий и других микроорганизмов оказалась столь замечательной, что, естественно, возникло представление об очень быстрой их фоссилизации. Это соображение было подтверждено серией лабораторных опытов по фоссилизации цианобактерий (Герасименко и др., 1994, 1996, 1999), которые показали, что фосфатизация при определенных условиях происходит всего за несколько часов. Такие же работы были проведены по изучению карбонатизации и окремнения (Westall et al., 1995; Бактериальная палеонтология, 2002), в ходе которых получен аналогичный результат.
Особенно важно выделить исследования по цианобактериальным матам (Заварзин, 1984, 1994; Герасименко, Заварзин, 1993) и биопленкам (в узком смысле).
В случае с цианобактериальными матами важны их древность, почти соизмеримая со всей историей Земли, их сложная организация, включающая совокупность разных типов бактерий и вертикальную зональность (Бактериальная палеонтология, 2002), удивительная их толерантность к широкому кругу условий и приуроченность к фотической зоне различных водоемов (от луж до океанов).
Что же касается более просто устроенных биопленок, то они распространены повсеместно, прежде всего на всех поверхностях и особенно там, где имеется любая степень смачиваемости. Их толщина может измеряться всего лишь первыми десятками нанометров. Основная масса биопленок состоит из полисахаридов, продуцируемых отдельными клетками обычно микронной размерности, но нередко и размером в первые сотни нанометров (так называемые нанобактерии) (Falk and Lynch, 1997). При этом фоссилизация тончайших биопленок тоже происходит стремительно, и они также могут хорошо сохраняться в ископаемом состоянии (рис. 2). В контексте настоящей статьи особенно следует подчеркнуть пронизанность рыхлых осадков, как морских, так и пресноводных, бактериями и полисахаридными пленками, играющими огромную роль как при фиксации осадка и сохранении следов, так и при диагенезе. Параллельно с фосфоритами были подвергнуты изучению высокоуглеродистые породы (Жмур и др., 1993, 1994, 1996), а затем и все остальные осадочные образования докембрия и фанерозоя (Бактериальная палеонтология, 2002; Rozanov et al., 1999).

Рис. 2. Современные (а) и ископаемые (б) биопленки. Толщина биопленок несколько десятков нанометров.
Основной вывод из всех этих работ таков: бактерии и биопленки сохраняются очень хорошо в ископаемом состоянии, они присутствуют во всех осадочных полродах, поскольку очень быстро фоссилизируются (обычно всего за несколько часов). Такое положение вещей вполне соответствует тому, что сегодня бактерии встречаются везде, в любых средах и на любых поверхностях, и без их участия не обходится ни выветривание, ни перенос материала, ни седиментация, ни диагенез осадков.
Как уже говорилось ранее (Розанов, 1999), еще в XIX в. было ясно, что образование железных и серных месторождений сопряжено с деятельностью бактерий. В начале и середине ХХ в. представление об огромной роли бактерий при формировании различных полезных ископаемых было очень распространено. Однако наши крупные ученые - академики Н.М. Страхов, А.В. Пейве и проф. Л.В. Пустовалов, решительно осудили такой образ мыслей, и долгое время разговоры на эту тему считались неприличными.
Однако, в связи с работами по бактериальному обогащению руд (Полькин и др., 1982; Karavaiko, Groudev, 1985) и работами по биоминералогии (Kirshvink et al., 1985; Lowenstam, Weiner, 1989; Rozanov, Barskov, 1998), значение бактерий при формировании минералов и пород обсуждалось неоднократно и породило множество интереснейших результатов. Более 20 лет назад Г. Ловенштам и С. Вейнер (Lowenstam, Weiner, 1989) указали в своей сводке около 20 минералов, образующихся при активном участии бактерий. Это могут быть образования внутри клеток, такие как магнетит или сульфиды железа у магнетотактических бактерий (Kirshvink et al., 1985; Devouagard et al., 1998; Posfai et al., 1998 ), в чехлах, как фосфатные и другие минералы (Герасименко и др., 1999; Zhegallo et al., 2000), а иногда одновременно и внутри, и снаружи клетки (Westall et al., 1995), проходя через оболочку, как в случае с марганцевыми минералами (Tebo et al., 1997). Несомненно, что маты и биопленки могут служить фильтром и таким образом накапливать те или иные элементы (Жмур и др., 1994). Несомненна также роль гликокаликса (или полисахаридов в пленках) как катализатора.
Сегодня описано уже более 100 минералов, образование которых может быть связано с деятельностью бактерий (Tazaki et al., 1997; Banfield and Nealson, 1997; American Mineralogist, 1998; Fortin et al., 1998; Герасименко и др., 1999). Особенно интересно обнаружение среди них кварца, кристобалита, барита, полевого шпата, слоистых силикатов (глинистых минералов), многочисленных минералов железа и марганца, доломита (Vasconcelos et al., 1995) и даже форстерита (Gorbushina et al., 2001). Получены многочисленные экспериментальные и природные подтверждения этому. Особенно впечатляют работы японских исследователей во главе с К. Тазаки (Tazaki et al., 1997; Tazaki, 1999). Результаты изучения ископаемых бактерий показывают, что минералообразование под влиянием бактерий или с их участием – процесс, который происходил на Земле всегда (Бактериальная палеонтология, 2002; Ponomarenko, 2002; Rozanov, 2002; Simakova, 2002).
Исследование бактериального образования минералов, особенно породообразующих, еще только начинается, и поэтому сегодня очень трудно определить пропорции терригенной и аутигенно-бактериальной составляющих, например, глинистых отложений. Еще сложнее решить вопрос с карбонатами, поскольку бактерий хорошей сохранности в этих породах почти нет, хотя очевидность биогенного, а часто и собственно бактериального, происхождения карбонатов, в том числе микратов, как правило, не вызывает сомнений (Buczynski, Chafetz, 1991; Kazmierczak et al., 1996). Анализ многочисленных опубликованных рентгено-структурных данных по бактериально образованным минералам показывает, как правило, наличие обильной аморфной фазы. Однако в случае с карбонатами мы этого не наблюдаем. Не исключено, что очень короткая аморфная стадия у арагонита и кальцита и быстрая кристаллизация этих минералов сразу разрушает клеточные оболочки и препятствует образованию объектов хорошей сохранности.
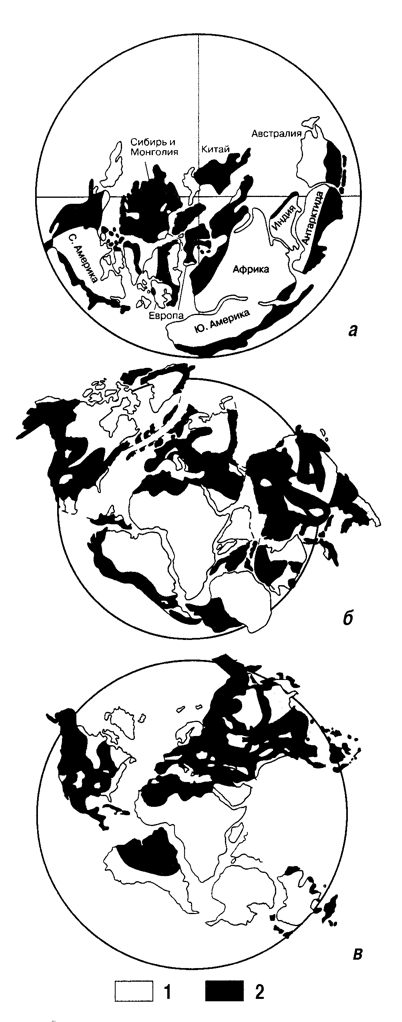
Рис. 3. Распределение эпиконтинентальных бассейнов в раннем кембрии (а), раннем ордовике (б) и позднем девоне (в) по Розанову, 1999. Обозначения: 1 – суша, 2 – море.
Теперь кратко остановимся на специфике эпиконтинентальных бассейнов прошлого. Прежде всего отметим, что их аналогов бассейнов в настощее время практически нет. Это были огромные по площади мелководные бассены, занимавшие часто большую часть кратонов (рис. 3). Дно этих бассейнов находилось преимущественно в пределах фотической зоны. Это приводило к тому, что вся водная масса и донные осадки были буквально напичканы бактериями или, точнее, микроорганизмами. Несмотря на малые глубины, придонные аноксические ситуации временами были обычным явлением. Чрезвычайно характерные линзовидные литологические тела небольшой мощности в разрезах платформенных областей, выклинивающиеся по простиранию и ассоциирующиеся также с выклинивающимися горизонтами палеопочв и аэрируемых поверхностей, являются показателем существования мигрирующих островов и систематического изменения донного рельефа. В отличие от эпиконтинентальных морей, современные океанические бассейны имеют насыщенный бактериальный мир только в приповерхностном и придонном слоях. В силу этих обстоятельств модели седиментации, разработанные для современных океанических ситуаций, непригодны для расшифровки седиментогенеза эпиконтинентальных бассейнов прошлого. Насыщенность водной массы микроорганизмами создавала трофическую специфику и специфику седиментогенеза. В частности, это касается организации транспорта на дно глинистого материала. Следует задуматься над тем, что пеллетные частицы – это не только продукт планктонных фильтраторов (зоопланктона), но и результат бактериальной деятельности, приводящей к образованию пеллетоподобных частиц. Насыщенность воды и осадка микроорганизмами и полисахаридами способствовала сохранению в ископаемом состоянии следов ряби и деликатных по строению организмов. Здесь же замечу, что присутствие волноприбойных знаков в докембрийских отложениях фиксирует наличие в это время в данной обстановке бактерий.
Для понимания истории становления жизни на Земле необходимо суммировать известные палеонтологические данные.
Должны быть рассмотрены три группы докембрийских фоссилий: хемофоссилии, или биомаркеры; собственно остатки организмов и ископаемые следы. Существует множество публикаций на эту тему. Однако хотелось бы выделить две крупные сводки В. Шопфа (Sсhopff, 1983; Schopf, Klein, 1992), в которых на то время были подведены итоги изучения микрофоссилий и строматолитов докембрия. Обобщенные результаты приведены в первой работе на фиг. 15-1. Ключевые моменты, отмеченные Шопфом и его соавторами, показаны на рис. 4. Кроме этого, следует отметить сборник под редакцией С. Бенгтсона (Bengtson, 1994) и статью А. Ноулла (Knoll, 1992).
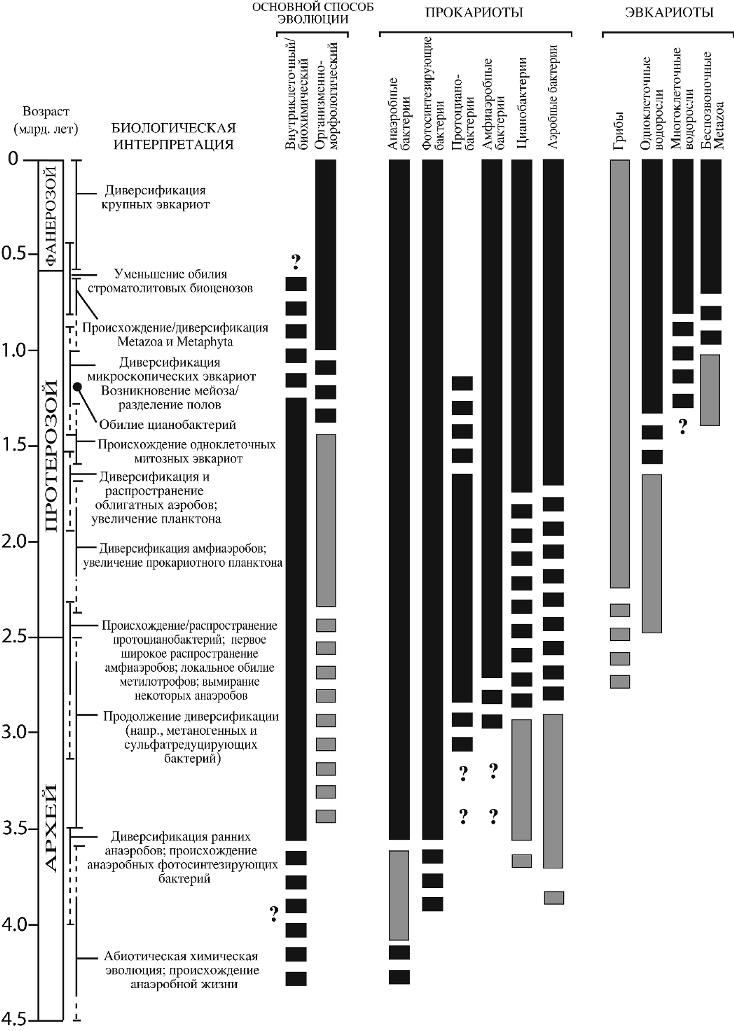
Рис. 4. События в эволюции органического мира (по Sсhopf, 1983, с дополнениями). Черное – по Шопфу, серое – наши дополнения.
Что же стало известно за последние 10-15 лет и насколько изменились наши знания и представления о начальных этапах развития жизни?
Биомаркеры. Интенсивное изучение хемофоссилий, или биомаркеров, привело к тому, что стали появляться данные о находках стеролов и о существовании эвкариот около 2.7 млрд. лет тому назад (Brooks et al., 1999). При этом количество кислорода в атмосфере должно было быть не менее 1 % от его количества в настоящее время. Как замечают в этой связи другие американские коллеги (Brake et al., 2002), появление в это время одноклеточных эвкариот само по себе должно было также повлиять на процесс увеличения кислорода и, соответственно, на окисление железа в водах морей.
Собственно остатки организмов. Среди этих фоссилий следует остановиться на трех разных группах: 1) остатках эвкариотных многоклеточных организмов, 2) остатках эвкариотных одноклеточных организмов, 3) остатках прокариот.
Остатки многоклеточных эвкариот. Идя вниз по стратиграфической шкале от широко известных ископаемых вендо-эдиакарского типа , прежде всего следует указать на рифейские аннелидоподобные организмы севера Европейской России и Китая (Weigou et al., 1986; Гниловская, 1998; Гниловская и др., 2000) и на Udokania problematica Leites, описанную много лет назад из удоканской серии Сибири. Поначалу не вызывавшая сомнений находка удоканий, по мнению В.В. Меннера, Б.С. Соколова, Р.Ф. Геккера и других, была сочтена за остатки либо кишечнополостных, либо кольчатых червей. Некоторые исследователи эти образования посчитали абиогенными, и они были выброшены из палеонтологической летописи. Однако никто не опубликовал никаких серьезных аргументов в пользу такой точки зрения. Специальное переизучение многочисленных дополнительных образцов позволило убедительно показать их биогенную природу. Т.А. Саютина и Е.С. Вильмова (1990) описали два вида Udokania и дали четко аргументировали невозможность считать эти образования псевдоморфозами по скаполиту, как это полагал Л.И. Салоп (1982). Но даже после этого зарубежные авторы обзоров и статей по эволюции жизни в докембрии (Bengtson et al., 1994; Knoll, 1992) и ряд российских исследователей игнорировали эти данные. Неудивительно, когда это делают англоязычные авторы, но плохое знание русской литературы российскими специалистами обескураживает.
Еще в конце 70-х – начале 80-х годов З. Валенчак (Walenczak, 1978, 1979, 1981) опубликовал первые результаты исследования дефектных структур и псевдоморфоз в древних породах методом темного поля под оптическим микроскопом. Выяснилось, что в метаморфических породах органическое вещество влияет на образование кристаллов, создавая дефектные структуры. Таким образом, была обнаружена возможность распознавания «теней» биологических объектов, находившихся в породе до момента перекристаллизации. Все эти работы были опубликованы на польском языке и остались незамеченными. Хотя позже краткая заметка Валенчака и моя в «Палеонтологическом журнале» была опубликована по-русски (Валенчак, Розанов, 1986) и переведена на английский язык, эта информация была вновь проигнорирована. В этой заметке описывались остатки возможных грибов из амфиболитов фундамента Восточно-Европейской платформы и графитовых мраморов слюдянской сериии Южного Прибайкалья.
Крайне интересны так называемые Grypania из протерозойских отложений Северной Америки и Китая (Han, Runnigar, 1992; Weigou, 1994). Обычно их находили в верхах протерозоя. Однако Хен и Раннигер описали грипаний из формации Негауни Айрон Мичигана, возраст которой ~ 2.1 млрд. лет. Похоже, большинство исследователей сходится на том, что грипании несомненно принадлежат многоклеточным эвкариотным организмам, вероятнее всего, водорослям.
Следующую группу ископаемых, относимых к эвкариотам, представляет ассоциация Chuaria-Tawuia. Особенно подробно этих ископаемыех исследовал Г. Хофман. Последняя его сводка опубликована в сборнике под редакцией С. Бенгтсона (Bengtson, 1994).
Одноклеточные эвкариоты. К числу вероятных одноклеточных эвкариотных организмов следует прежде всего отнести разнообразные акритархи. Можно достаточно уверенно полагать их присутствие уже в ганфлинтской биоте (Sсhopff, 1983; Knoll, 1994; Бактериальная палеонтология, 2002).
Прокариоты. О находках прокариот я уже писал выше. Но все же напомню достаточно интересные данные из раннего архея (Walsh, 1992; Schopf, 1993; Весталл, Велш, 2002). В раннем проторозое количество находок очень велико (Sсhopf 1983; Бактериальная палеонтология, 2002 и т.д.). Рифей же просто напичкан остатками прокариот. Однако наибольший интерес представляют находки, достаточно обоснованно интерпретируемые как цианобактерии. Мне представляется, учитывая время появления кислорода, что первые же находки строматолитов можно смело считать временем достоверного появления цианобактерий.
Ископаемые следы. В обширной литературе по этому вопросу главное внимание обращалось на изменение характера ископаемых следов при переходе от докембрия к кембрию. Действительно, картина распределения следов в докембрии и кембрии достаточно контрастная. Эта резкая разница привела к тому, что целый ряд исследователей пришел к выводу о возможности использования следов для фиксации границы кембрия и докембрия. Мы уже писали о фатальной ошибке в этих рассуждениях (Розанов и др., 1997), в результате чего выбранная граница стала неопределимой за пределами типового района на Ньюфаундленде. Однако применительно к данной статье мне представляется, что увлечение данным пограничным переходом привело к снижению внимания к докембрийским следам и особенно к их поиску в архейских и нижнепротерозойских отложениях. Вместе с тем, совсем недавно были описаны интереснейшие следы (возможно, слепки), названные Horodyskia и интерпретированные как следы колониальных тканевых организмов (Fedonkin, Yochelson, 2002; Gray et al., 2002). Возраст этих следов ~ 1.5 млрд. лет. Еще ранее были упомянуты следы, обнаруженные Б. Шишкиным (Новосибирск) на Сибирской платформе (Игарский район) в чернореченской свите нижнего рифея. По своей форме (и, следовательно, характеру движения) они несомненно принадлежат целоматным червеобразным организмам, а некоторые из них, возможно, даже артроподам.
Таким образом, на основе палеонтологических данных намечается весьма логичная последовательность появления (обнаружения) разных групп организмов: бактерии ~ 3.8-4.0 млрд. лет (при этом, вероятно, правильно, что вначале появились или начали широко функционировать анаэробные бактерии и несколько позднее аэробные), цианобактерии ~ 3.5-3.8 млрд. лет, одноклеточные эвкариоты ~ как минимум 2.7 млрд. лет, грибы не позднее ~ 2.4 млрд. лет (а вероятно, древнее), многоклеточные водоросли и низшие Metazoa (ацеломаты) древнее ~ 2.1 млрд. лет) а целоматы древнее ~ 1.5-1.6 млрд. лет.
С большой долей уверенности можно считать, что наша биосфера, как полагают многие, сначала была прокариотной. Затем очень продолжительное время она находилась в «промежуточном» состоянии, когда постепенно появлялись прокариоты (внутри этого периода можно наметить более мелкие этапы, например, появление целомат и т.д.). И, наконец, в фанерозое биосфера приобретает современный вид, когда решающую роль уже играют эвкариоты.
Теперь несколько слов о составе древней атмосферы и времени появления кислорода. В.И. Виноградов (1980) на основе исследований изотопов серы уже довольно давно высказал предположение, что кислородная атмосфера, сходная с современной, существует практически с архея. Эти представления были встречены, мягко говоря, прохладно, и до сих пор существует устойчивое убеждение, что в архее атмосфера долгое время была восстановительной.
Данные, которые обсуждатся в настоящей статье, недвусмысленно указывают на достаточно раннюю оксигенацию атмосферы. Ранее уже говорилось о стеролах с возрастом 2.7 млрд. лет, свидетельствующих о наличии в этом времени эвкариот. Добавив сюда грибы из амфиболитов фундамента Восточно-Европейской платформы и графитовых сланцев Сибири, удоканий, вероятных кишечнополостных, следы целоматных животных и эвкариотный планктон (акритархи) из ганфлинта и его аналогов, получим совершенно иную картину (рис. 5). К выводам такого толка, исходя из самых разных фактов, приходят сейчас многие исследователи. Интересны, например, исследования Я. Ватанабе и Х. Омото (Watanabe, Ohmoto, 2002) цериевых аномалий из древних палеопочв Южной Африки, возраст которых составляет 2.4-2.6 млрд. лет. Эти авторы настаивают на существовании в то время кислородной атмосферы.
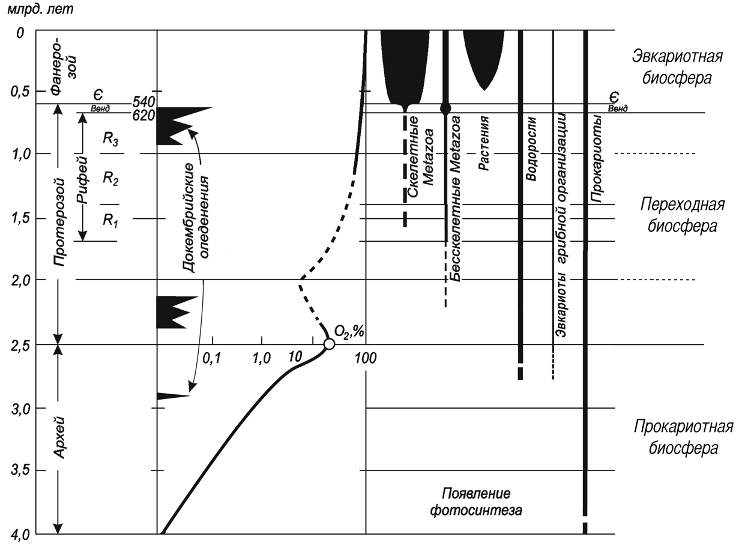
Рис. 5. Появление разных групп организмов в истории Земли и схема оксигенезации атмосферы.
Очень важные результаты были получены и по балансу урана в архейском “океане” (Yamaguchi, Ohmoto, 2002). Я же обратил внимание на недостаточную корректность примера с уранинитом, приведенного группой авторов в книге Шопфа (Schopf, 1983) (глава 2, рис. 11-13). Изображенные там зерна уранинита не могут быть окатанными обломочными зернами в силу того, что они слишком малы (менее 0.15 мм). Я думаю, что необходимо внимательное переизучение этого материала. Не исключено, что форма зерен объясняется, скорее всего, их бактериальной природой. Таким образом, один из главных постулатов гипотезы восстановительной атмосферы того времени может оказаться несостоятельным. Можно еще и еще приводить примеры исследований, результаты которых говорят о раннем появлении кислорода в атмосфере. Приведу еще только один пример. Из архейских пород Канады (~ 2.7 млрд. лет) исследовались изотопы кислорода в гематитовых кристаллах. Полученные результаты позволяют считать, что в это время атмосфера была кислородной (Ikemi et al., 2002).
Гораздо более ранняя, чем обычно считалось, оксигенация атмосферы подкрепляет представление о достаточно раннем появлении цианобактерий и цианобактериальных матов. Во всяком случае, не исключено, что ископаемые Шопфа из Австралии, и Велш и Весталл из Южной Африки могут оказаться цианобактериями (3.5 млрд. лет), тем более, что Весталл описала ископаемых бактерий и стромалитоподобные образования.
Я не стану обсуждать результаты исследования углистых хондритов, но в свете сказанного выше и здесь есть над чем задуматься (Gerasimenko et al., 1999).
Хотел бы сердечно поблагодарить за помощь моих коллег: В.Ю. Дмитриева, М.К. Емельянову, Е.А. Жегалло, Н.В. Леонову, М.А. Федонкина, Ю.Я. Шабанова и А.А. Шевырева. Исследования были поддержаны грантами РФФИ (проект № 02-04-48094) и НШ (№ 974.2003.5).
Бактериальная палеонтология. М.: ПИН РАН, 2002. 188 с.
Валенчак З., Розанов А.Ю. Древнейшие эвкариоты? // Палеонтол. журн. 1986. № 4. С. 99-101.
Весталл Ф., Велш М. Ископаемые бактерии и бактериальные биопленки // Бактериальная палеонтология. М.: ПИН РАН, 2002. С. 84-90.
Виноградов В.И. Роль осадочного цикла и геохимии изотопов серы. М.: Наука, 1980.
Герасименко Л.М., Гончарова И.В., Жегалло Е.А. и др. Процесс минерализации (фосфатизации) нитчатых цианобактерий // Литология и полезн. ископаемые. 1996. № 2. С. 208-214.
Герасименко Л.М., Гончарова И.В., Заварзин Г.А. и др. Динамика высвобождения и поглощения фосфора цианобактериями // Экосистемные перестройки и эволюция биосферы. М.: Недра, 1994. Вып. 1. С. 348-458.
Герасименко Л.М., Заварзин Г.А. Реликтовые цианобактериальные сообщества // Проблемы доантропогенной эволюции биосферы. М.: Наука, 1993. С. 222-254.
Герасименко Л.М., Заварзин Г.А., Розанов А.Ю., Ушатинская Г.Т. Роль цианобактерий в образовании фосфатных минералов // Журн. общ. биол. 1999. Т. 60. № 4. С. 415-430.
Гниловская М.Б. Древнейшие аннелидоморфы из верхнего рифея Тимана // Докл. РАН. 1998. Т. 359. № 3. С. 369-372.
Гниловская М.Б., Вейс А.Ф., Беккер Ю.Р. и др. Доэдиакарская фауна Тимана (аннелидоморфы верхнего рифея) // Стратиграфия. Геол. корреляция. 2000. Т. 8. № 4. С. 11-39.
Жмур С.И., Горленко В.М., Розанов А.Ю. и др. Цианобактериальная бентосная система – продуцент углеродистого вещества шунгитов нижнего протерозоя Карелии // Литология и полезн. ископаемые. 1993. № 2. С. 122-124.
Жмур С.И., Розанов А.Ю., Кравченко С.М., Жегалло Е.А. О генезисе богатых редкоземельно-ниобиевых руд Томтора (север Сибирской платформы) // Докл. РАН. 1994. Т. 336. № 3. С. 372-375.
Жмур С.И., Розанов А.Ю., Лобзова Р.В., Жегалло Е.А. Об источнике углерода графитовых руд Ботогольского щелочного массива (Восточный Саян) // Докл. РАН. 1996. Т. 348. № 3. С. 360-362.
Заварзин Г.А. Бактерии и состав атмосферы. М.: Наука, 1984. 199 с.
Заварзин Г.А. Реликтовые прокариотные сообщества гипергалинных водоемов морского происхождения // Экосистемные перестройки и эволюция биосферы. М.: Недра, 1994. Вып. 1.
Полькин Г.И., Адамов Э.В., Панин В.В. Технология бактериального выщелачивания цветных и редких металлов. М.: Недра, 1982. 288 с.
Розанов А.Ю. Ископаемые бактерии и новый взгляд на процесс седиментации // Соросовский образовательный журн. 1999. № 10. С. 63-68.
Розанов А.Ю., Миссаржевский В.В., Волкова Н.В. и др. Томмотский ярус и проблема нижней границы кембрия. М.: Наука, 1969. Труды ГИН АН СССР. Т. 206. 380 с.
Розанов А.Ю., Жегалло Е.А. К проблеме генезиса древних фосфоритов Азии // Литология и полезн. ископаемые. 1989. № 3. С. 67-82.
Розанов А.Ю., Заварзин Г.А. Бактериальная палеонтология // Вестник РАН. 1997. Т. 67. № 3. С. 241-245.
Розанов А.Ю., Семихатов М.А., Соколов Б.С. и др. Решение о выборе стратиграфической границы докембрия и кембрия: прорыв в проблеме или ошибка? // Стратиграфия. Геол. корреляция. 1997. Т. 5. № 1. С. 21-31.
Салоп Л.И. Геологическое развитие Земли в докембрии. Л.: Недра, 1982. 343 с.
Саютина Т.А., Вильмова Е.С. Проблематичные ископаемые Udokania Leites из протерозойских отложений Забайкалья // Палеонтол. журн. 1990. № 3. С. 100-105.
American Mineralogist. 1998. V. 83. N 11-12 (2). P. 1387-1607.
Banfield J.F., Nealson K.H. (eds.) Geomicrobiology: Interactions between microbes and minerals // Rev. Mineralogy. 1997. V. 35. Р. 1-448.
Barghoorn T.S., Tylor S.A. Microorganisms from the Guntflint cherts // Science. 1965. V. 147. N 3658. P. 563-577.
Bengtson S. (ed.) Early life on Earth. N. Y. Columbia Univ. Press, 1994. 630 p.
Brake S.S., Hasiotis S.T., Dannally H.K., Connors K.A. Eukariotic stromatolite builders in acid mine drainage: Implications for Precambrian iron formations and oxygenation of the atmosphere? // Geology. 2002. V. 30. N 7. P. 599-602.
Brooks J.J., Logan G.A., Buick R., Summons R.E. Archean molecular fossils and the early rise of eukaryotes // Science. 1999. V. 285. P. 1033-1036.
Buczynski C., Chafetz H.C. Habit of bacterially induced precipitales of calcium carbonate and the influence of medium visocity on mineralogy // J. Sediment. Petrology. 1991. V. 61. P. 226-233.
Devouagard B., Posfai M., Xin Hua et al. Magnetite from magnetotactic bacteria: Size distributions and twinning // Amer. Mineralogist. 1998. V. 83. Р. 1387-1398.
Fedonkin M.A., Yochelson E.L. Middle Proterozoic (1.5 Ga) Horodyskia moniliformis Yochelson and Fedonkin, the oldest known tissue-grade сolonial еucaryote // Smithson. Contrib. Paleobiology. 2002. N 94. Р. 1-29.
Folk R.L., Lynch F.L. The possible role of nannobacteria (dwart bacteria) in clay mineral diagenesis and the importance of careful sample preparation in high magnification SEM stady // J. Sedimentary Res. 1997. V. 67. P. 597-603.
Fortin D., Ferris F.G., Scott S.D. Formation of Fe-silicates and Fe-oxides on bacterial surfaces in samples collected near hydrothermal vents on the Southern Explorer Ridge in the northeast Pacific Ocean // Amer. Mineralogist. 1998. V. 83. N 11-12 (2). P. 1399-1408.
Gerasimenko L.M., Hoover R.B., Rozanov A.Yu. et al. Bacterial рaleontology and studies of сarbonaceous сhondrites // Paleontol. J. 1999. N 4. P. 439-459.
Gorbushina A., Boettcher M., Brumsack H.-J. et al. Biogenic forsterite and оpal as a product of biodeterioration and Lichen Stromatolite Formation in Table Mountain Systems (Tepuis) of Venezuela // Geomicrob. J. 2001. N 18. P. 117-132.
Gray K., Williams I.R., Martin D.M.B. et al. New occurrences of «strings of beads” in the Bangemall Supergroup: a potential biostratigraphic marker horizon // Ann. Rep. Geol. Surv. West. Australia. 2002. P. 69-73.
Han T.M., Runnigar B. Megascopic eukaryotic algae from the 2.1-billion-year-old Negaunee Iron Formation, Michigan // Science. 1992. V. 257. P. 232-235.
Ikemi H., Ohmoto H., Nakamura T., Shimada N. The оrigin of hematite сrystals associated with Archean redbeds in the Lake Shebandowan Area, Canada: A preliminary study from сhemical сomposition and оxygen isotopes // Astrobiology. 2002. V. 2. N 4. P. 562.
Karavaiko G.I., Groudev S.N. (eds.) Biogeotechnology of metals // Proceed. Intern. Seminar and Intern. Training Course. Moscow, 1985. 417 р.
Kazmierczak J., Colemen M.L., Gruszczynski M. et al. Cyanobacterial key to the genesis of micritic and peloidal limestones in ancient seas // Acta paleontol. рolon. 1996. V. 41. P. 319-338.
Kirshvink J.L., Jones D.S., MacFadden B.J. (eds.). Magnetite biomineralization and magnetoreception in оrganisms. N. Y.; L.: Plenum, 1985. V. 2.
Knoll A.H. The early evolution of eukaryotes: A geological perspective // Science. 1992. V. 256. N 5057. P. 622-627.
Knoll A.H. Neoproterozoic evolution and end nomental change // Early life on Earth. N. Y.: Columbia Univ. Press, 1994. P. 439-449.
Lowenstam H.A., Weiner S. 0n biomineralization. Oxford: Oxford Univ. Press, 1989. 324 p.
Ponomarenko A.G. Bacteriomorph structures in Mesozoic lake deposits // Proceed. SPIE. 2002. V. 4939. P. 88-92.
Posfai M., Buseck P., Bazylinski D., Frankel R. Iron sulfides from magnetotactic bacteria: Structure, composition and phase transitions // Amer. Mineralogist. 1998. V. 83. N 11-12. Pt. 2. P. 1469-1481.
Rozanov A.Yu. Some problems of bacterial mineralization and sedimentation // Proc. SPIE. 2002. V. 4939. Р. 83-87.
Rozanov A.Yu., Barskov I.S. Diversity and phylum distribution of biominerals // Advanced Mineralogy. 1998. V. 3. P. 247-255.
Rozanov A.Yu., Zhegallo E.A., Hoover R. Microbiota of the Botogol graphites // Proceed. SPIE. 1999. N 3755. P. 38-46.
Schopf J.W. (ed.) Earth’s еarliest biosphere: Its оrigin and еvolution. Princeton: Princeton Univ. Press, 1983. 544 p.
Schopf J.W. Microfossils of the Early Archean Apex Chert: New evidence of the antiquity of life // Science. 1993. V. 260. N 5180. P. 640-646.
Schopf J.W., Klein C. (eds.). The Proterozoic biosphaere: А multidisciplinary study // N. Y.: Cambridge Univ. Press, 1992. 1348 p.
Simakova Y.S. Proposed role of bacteria in volkonskoite formation // Proceed. SPIE, 2002. V. 4939. P. 117-124.
Tazaki K. Architecture of biomats reveals history of geo-, aqua- and biosystems // Episodes. 1999. V. 22. N 1. P. 21-25.
Tazaki K., Aoki A., Asada R., et al. A new world in the science of biomineralization - еnviromental biomineralization in microbial mats in Japan // Sci. Reports Kanazawa Univ. 1997. V. 42. N 1-2. P. 1-64.
Tebo B.M., Ghiorse W.C., Van Waasbergen L.G. et al. Bacterially mediated mineral formation: Insights into manganese (II) oxidation from molecular genetic and biochemical studies // Rev. Mineralogy. 1997. V. 35. P. 225-266.
Vasconcelos C., McKeuzi J.A., Bernasconi S. et al. Microbial mediation as a possible mechanism for natural dolomite formation at low temperature // Nature. 1995. V. 377. P. 220-222.
Walenczak Z. Metody defectoskopii optycznej w badaniach from organicznego pochodzenia w skalach // Przegl. Geol. Warszawa. 1978. N 3. P. 172-175.
Walenczak Z. Drobnodyspesyjna forma wyste, powania substancii organiczen w skalach // Biul. Inst. Geol. 1979. N 315. P. 151-166.
Walenczak Z. Wlasciwosci pamieciowe drobnodyspersyjnych domieszek w mineralach i mozliwosci ich wykorzystania // Biul. Inst. Geol. 1981. N 334. P. 5-58.
Walsh M.M. Microfossils and possible microfossils from the Early Archean Onverwacht Group, Barbeton Mountain Land, South Africa // Precambrian Res. 1992. V. 54. P. 271-293.
Watanabe Y., Ohmoto H. Ce-anomalies in the 2.6-2.4 Ga Kalkkloof paleosol in S. Africa: Evidence for the early development of an oxygenated atmosphere // Astrobiology. 2002. V. 2. N 4. P. 585.
Weigou Sun, Guixiang Wang, Benhe Zhou. Macroscopic wormlike body fossils from Upper Precambrian (900-700 Ma), Huainan district, Anhui, China and their stratigraphic and evolutionary significance // Precambrian Res. 1986. V. 31. P. 377-403.
Weigou S. Early multicellar fossils // Early life on Earth. N. Y.: Columbia Univ. Press, 1994. N 84. P. 358-369.
Westall F., Boni L., Guerzoni E. The experimental siliсification of microorganisms // Palаeontology. 1995. V. 38. Pt. 3. P. 495-528.
Yamaguchi K., Ohmoto H. Uranium budget of the Archean оceans: Implications for the Redox еvolution of the аtmosphere // Astrobiology. 2002. V. 2. N 4. P. 585-586.
Zhegallo E.A., Rozanov A.Yu., Ushatinskaya G.N. et al. Atlas of microorganisms from ancient phosphorites of Khubsugul (Mongolia). Huntsville, NASA, 2000. 167 p.