
Проблемы Эволюции |
1968
Теория стабилизирующего отбора
I. ИНДИВИДУАЛЬНАЯ ИЗМЕНЧИВОСТЬ КАК ОСНОВА ИСТОРИЧЕСКОЙ ИЗМЕНЯЕМОСТИ ОРГАНИЗМОВ
Особи любого вида организмов — как растений, так и животных — отличаются друг от друга множеством индивидуальных особенностей. Анализ этих различий позволяет вскрыть некоторые закономерности в их распределении среди особей, происходящих от определенных родоначальных форм, а также среди особей, живущих в определенных условиях внешней среды или в определенном сезоне и т. п.
Экспериментальный анализ позволяет проникнуть глубже в существо этих различий: одни из них, возникнув у отдельной особи, встречаются затем примерно в том же выражении в потомстве (у большего или меньшего числа особей); другие, проявляясь в известных условиях у всех особей, исчезают у потомства (сразу или постепенно), если это последнее развивается в других условиях внешней среды.
В первом случае мы лишь предполагаем существование какого-то влияния факторов внешней среды на организм и на его половые клетки. Если рассматривать такое изменение организма (половой клетки), как его реакцию, то — во всяком случае, до сих пор,— оказалось невозможным установить явную связь между факторами внешней среды и специфической наследственной реакцией организма (половой клетки). Дарвин считал, что эта специфика определяется в основном индивидуальными свойствами каждой особи и называл поэтому такие изменения индивидуальными и «неопределенными». Теперь их называют мутациями. Внешнее выражение мутаций зависит, во всяком случае, от индивидуальных наследственных свойств самого организма.
Во втором случае с легкостью устанавливается связь между определенными факторами внешней среды и характером изменения организма. Специфика реакции явно определяется в основном самим организмом, и Дарвин назвал такие изменения массовыми или, очень удачно, «определенными» изменениями. Теперь их называют модификациями. Их выражение безусловно зависит от наследственных свойств организма, однако не столько от его индивидуальных свойств (что все же имеет значение), сколько от общих наследственных свойств организма как представителя известного вида. Обычные модификации имеют приспособительный характер и закономерно повторяются у различных особей данного вида организмов. Как и другие их свойства, как и вся организация вообще (вместе с ее функциями), способность к определенным приспособительным модификациям есть продукт длительного исторического развития организмов в разнообразных, но все же определенных условиях внешней среды.
1. РОЛЬ ВНЕШНИХ И ВНУТРЕННИХ ФАКТОРОВ В ВОЗНИКНОВЕНИИ ИЗМЕНЕНИЙ
Как в мутациях, так и в модификациях мы имеем изменения организмов, зависящие — в своем возникновении и в своем выражении — от наследственных свойств организма и от факторов внешней среды. Специфика изменений в обоих случаях зависит от наследственных свойств самого организма.
При развитии любой особи факторы внешней среды выступают в основном лишь в роли агентов, освобождающих течение известных формообразовательных процессов и условий, позволяющих завершить их реализацию. И это мы должны рассматривать как результат исторического развития организма в определенных условиях внешней среды. В историческом развитии организма менялись формы взаимодействия между организмом и средой. Эволюция шла в общем под знаком освобождения развивающегося организма из-под власти случайных явлений во внешней среде.
Это сопровождалось развитием внутренних регуляторных механизмов, контролирующих процессы индивидуального развития. Освобождение организма от детерминирующей роли факторов среды именно и означает установление системы внутренних факторов развития, определяющих специфическое течение формообразовательных процессов.
Если на начальных этапах своего возникновения живые существа были в полной власти случайных изменений внешних факторов, то все их дальнейшее развитие состояло в постепенном освобождении от этой зависимости. Оно выражалось в их самоопределении как некоторых, до известной степени свободных, носителей жизни, обладавших своей формой (величиной, структурой) и своими функциями. Влияния внешних факторов ближайшего окружения вводились постепенно в определенное русло. Они преобразовывались в организме вследствие его специфических ответных реакций. Против неблагоприятных влияний организм вырабатывал средства пассивной или активной защиты. Благоприятные влияния использовались организмом; ответные изменения вводились в определенное русло наиболее выгодных форм реагирования. Это было связано с дифференцированием организма. Каждая дифференцировка означает использование организмом определенных связей со средой. Таким образом, различные факторы внешней среды все более осваиваются организмом. Организм теперь сам определяет свое отношение к внешним факторам, защищаясь от одних влияний и используя другие. Это отношение к внешней среде оказывается для разных организмов весьма различным. И формы защиты различны и использование факторов среды далеко не одинаково. Каждый вид организмов использует внешнюю среду по-своему и по-разному отвечает на изменения факторов внешней среды. Реакции организма на «освоенные» факторы среды всегда строго специфичны, и эта специфика определяется именно исторически обоснованными свойствами самого организма, его эволюцией в определенной среде, при непрерывном взаимодействии с данными его факторами. С другой стороны, не освоенные организмом факторы, если они вообще достигают организма и влияют на него, могут оказать лишь неопределенное воздействие, которое проявится в более или менее глубоком нарушении нормального строения и нормальных функций организма.
Результаты «освоения» факторов внешней среды сказываются, следовательно, в защитных реакциях организма против вредных воздействий и в использовании благоприятных факторов. В обоих случаях реакции могут иметь характер легко и быстро обратимых физиологических процессов или более медленно обратимых формообразовательных реакций. В последнем случае говорят о приспособительных модификациях или морфологических адаптациях.
«Неосвоенные» факторы внешней среды, если они достигают организма и могут оказать на него влияние, не встречают в организме подготовленной базы. Организм исторически развивался вне взаимодействия с данными факторами (например — лучи рентгена, ультрафиолетовые лучи значительной концентрации, ультразвуковые колебания, различные необычные химические вещества, попавшие в состав пищи или окружающей водной или воздушной среды) или с данными их интенсивностями (просто необычные для данного организма температуры, влажность, реакция среды, концентрация солей и многие другие). Поэтому организм не выработал против них ни средств защиты, ни, тем более, средств их использования. Беззащитный организм не может ответить на такие воздействия определенным целесообразным изменением. Влияние этих факторов будет определяться, во-первых, качеством и интенсивностью самого воздействия и, во-вторых, индивидуальными свойствами самого организма (т. е. не просто его строением, но и, в особенности, его физиологическим состоянием). Такое влияние может быть только неопределенным. Неопределенными будут, следовательно, все новые изменения организма, не имеющие еще своего исторического прошлого. В эту категорию изменений войдут, однако, не только мутации, как новые «наследственные» изменения, но и любые новые, т. е. впервые возникающие, модификации. Такие модификации не имеют приспособительного характера, и их называют обычно морфозами. Морфозы могут быть двоякого происхождения: с одной стороны, внешний фактор, оказавший модифицирующее влияние, может быть «новым» (рентген, необычные химические влияния и т. п.), с другой стороны, сам организм может быть до известной степени «новым», т. е. измененным свежей мутацией. И в последнем случае модификации, касающиеся как раз выражения данной мутации, будут иметь характер морфозов.
В мутациях и в их модификациях мы имеем более или менее глубокие изменения организма и его реакций, определяемые взаимодействием организма с факторами внешней среды. В обоих случаях специфика изменения зависит в основном от свойств самого организма. Однако, как упомянуто в самом начале, мутация, раз возникнув, проявляется затем и у некоторых потомков измененной особи даже при иных условиях внешней среды, а модификация зависит в своем выражении всегда от определенных факторов внешней среды, при изменении которых эта модификация не повторяется ни у одной особи. Мутация есть всегда новоприобре-тение организма, а модификация есть некоторая надстройка — вариант существующей уже организации. Мутация передается потомству в строго закономерном порядке. Эти закономерности были вскрыты Г. Менделем. Они были в основном подтверждены и подверглись очень глубокому анализу в современной генетике (в особенности — в школе Т. Г. Моргана). Особенно следует подчеркнуть явления расщепления, указывающие на дискретность этого наследования, на несмешиваемость признаков в потомстве.
Модификации, как обыкновенно выражаются, ненаследственны. Это неточно и требует разъяснения. Сама способность к определенным модификациям вполне наследственна. С другой стороны, вся организация — как типичная, так и уклоняющаяся (в том числе и мутация) — не дана заранее, а развивается на известной наследственной основе (генотипе). Это развитие определяется, однако, не только наследственной основой, но и внешними факторами. Поэтому как генотипически «нормальный» организм, так и мутация получают в разных условиях внешней среды различное выражение. Эти различия в выражении нормальной организации или определенной мутации, зависящие в своем развитии от различных факторов внешней среды, и называют модификациями. Точно так же и процесс, ведущий к соответствующему изменению типичной организации или типичного выражения мутации, называют модификацией.
Модификация есть, следовательно, известная индивидуальная (вместе с тем исторически обоснованная) надстройка над существующей организацией, определяемая спецификой реакций данного организма на изменения в факторах внешней среды. Модификация определяется всегда и внешними факторами, но касается только процессов индивидуального развития, зависящих от этих факторов. Внешняя среда выступает здесь в роли факторов зависимого развития, определяющих (более или менее непосредственно или через функции организма) до известной степени качество его результата. В основном, однако, качество результата определяется именно наследственной основой данной организации, ее способностью к определенным модификациям. Поэтому генетики и говорят о наследственной «норме реакций». Для всех зависимых реакций характерна именно их определенная зависимость от внешних факторов в процессах индивидуального развития. Так как база этих реакций определяется наследственной основой, то она и в потомстве реализуется всегда тем же самым путем, т. е. требует для своего осуществления наличия тех же внешних факторов. Без этого данная реакция (и ее результат — модификация) не повторяется.
Некоторое влияние результатов индивидуальных реакций на развитие потомства, впрочем, иногда наблюдается: это — довольно редкие случаи длительных модификаций. Однако, в отличие от мутаций, длительные модификации передаются при половом размножении, во-первых, всему потомству, а не определенному проценту особей, во-вторых, в слитном, а не в дискретном проявлении, и, в-третьих, в быстро затухающем от поколения к поколению выражении. У нас имеются все данные связывать некоторую возможность длительных модификаций с наследственной передачей через плазму половых клеток. С другой стороны, дискретность наследования мутаций определяется строением ядерной субстанции (хромосом) . В случаях наследования длительных модификаций проявляется некоторая инертность живого вещества. В других случаях яснее выступает обратимость его реакций. Все морфофизиологиче-ские реакции организма до известной степени обратимы. Некоторые из них характеризуются очень быстрой обратимостью, Таково, например, явление мышечного сокращения и другие типично физиологические реакции, которые иногда связаны с явным изменением формы (как раз пример мышцы показывает эту связь) или с более тонким изменением структуры (железы, ретина глаза и т. п.). Другие реакции оказываются медленно обратимыми или необратимыми в пределах органа или особи. Таково большинство морфогенетических реакций (уменьшение поверхности листовой пластинки на свету; удлинение стебля в темноте и в тепле; удлинение волос млекопитающего при низкой температуре). Такие реакции мы и называем модификациями. Они очень разнообразны как по ширине охвата организации, так и по степени их обратимости.
Во всяком случае, нам следует подчеркнуть, что вся организация вместе со всеми ее реакциями имеет историческое обоснование, и это дает единственное объяснение определенности и адаптивности большинства реакций, в том числе и модификаций. Только в нарушениях исторически создавшейся наследственной основы выявляется неопределенность изменений организации (мутации) и неадаптивность ее реакций (морфозов). Мутации и их различные выражения не имеют еще исторического прошлого. Только в них мы имеем новые свойства организма, которые в известных случаях могут быть использованы в процессе исторических его преобразований. Интересно, что именно новощшобретения организма наследственны в полной мере (мутации). На мутациях строятся новые формы реагирования и, следовательно, не только новые признаки организации, но и различные ее выражения в различных условиях внешней среды, т. е. различные модификации.
2. МОДИФИКАЦИИ, ИХ ВЫРАЖЕНИЯ И ОБРАТИМОСТЬ
Иногда модификация касается отдельных признаков или органов. Так, например, в зависимости от цвета окружающего фона меняется окраска у молодых саламандр (Salamandra maculosa, пo Каммереру).
В зависимости от интенсивности освещения меняются величина, форма и структура листьев у многих растений.
В других случаях модификация касается в большей или меньшей степени всего организма. Различия в питании личинок приводят у насекомых обычно к весьма значительным различиям в величине взрослого насекомого.
Иногда способность к модификациям выражается в явной перестройке всей организации. Изменения имеют характер целостных и притом безусловно приспособительных преобразований. Организация оказывается полиморфной в зависимости от разных условий среды. Можно сказать, что такой организм обладает не одной нормой, а несколькими вполне законченными «адаптивными нормами». При ясно характеризуемых экологических различиях получаются «экофены» Турессона. Таковы различия между долинными и горными модификациями ряда растений.
Иногда разные адаптивные нормы отвечают различным сезонам (сезонный полиморфизм и цикломорфоз). Реже такой полиморфизм связывается с половыми различиями (например, у червя Bonellia) или сочетается с разделением труда внутри колонии, как у пчел и, в особенности, у муравьев и термитов.
Модификации различаются и по форме реагирования. Иногда реакция имеет характер непосредственного ответа на данный раздражитель, который прямо влияет на течение известных физиологических процессов. Таково, очевидно, непосредственное влияние температуры на процессы пигментации. Возможно, что и влияние температуры на скорость течения некоторых формообразовательных процессов, на скорость роста и скорость гистологической дифференцировки имеет иногда также характер непосредственного влияния на развивающиеся ткани организма. Однако в большинстве случаев, несомненно, сказывается весьма косвенное влияние факторов через более или менее сложную цепь зависящих друг от друга физиологических процессов. Самый характер зависимости дает нередко ясные на это указания. Таковы как раз все более сложные реакции, имеющие ясно выраженный адаптивный характер. Здесь нередко реагирующей тканью является вовсе не та ткань, которая воспринимает раздражение от внешнего фактора (обычно это эмбриональные и меристематические ткани, хорошо защищенные от различных внешних влияний). С другой стороны, и внешний фактор, вызывающий данную морфогенетическую реакцию, может быть совершенно не тем фактором, к которому организм приспособляется. Наконец, может быть и очень большое расхождение между временем восприятия известных раздражений и временем, когда это влияние обнаруживается в морфогенетиче-ской реакции. Так, у некоторых деревьев световая и теневая формы листьев предопределяются условиями освещения еще в течение предшествующего лета, а не весной, когда эти почки начинают развиваться.
При этом обычно нет никакой пропорциональности между интенсивностью внешнего фактора и количественным выражением модификации. В известных условиях минимальной интенсивности внешнего фактора, отвечающей нижнему порогу раздражимости тканей организма, сразу наступает типичная реакция в полном своем выражении. При дальнейшем увеличении интенсивности раздражителя до максимально допустимого уровня, отвечающего верхнему порогу раздражимости тканей, характер реакции организма не меняется. Все это указывает на существование физиологических корреляций, связывающих в организме ткани, воспринимающие раздражение и реагирующие, и на существование регуляторных механизмов, обеспечивающих не только наступление согласованной реакции различных частей организма, но и типичное их оформление именно при различных интенсивностях внешних факторов.
Хотя процессы индивидуального развития и зависят от известных внешних факторов, однако в этом взаимодействии между развивающимся организмом и средой лишь решается вопрос о включении известной, типичной для организма (в данных условиях) формообразовательной реакции. После некоторого, иногда очень краткого, периода «индукции», когда процесс «детерминируется», дальнейшее его течение идет автономно и без наличия этих определяющих факторов. Такое развитие можно назвать «авторегуляторным» (Шмальгаузен, 1940). Большая или меньшая степень «авторегуляции» характерна для всех адаптивных модификаций и тем более для всех вообще «нормальных» формообразовательных процессов.
Косвенный характер связи между внешними факторами и организмом обнаруживается еще резче в тех случаях, когда она осуществляется лишь через функции организма, которые меняются в особенности в связи с изменением поведения животного в той или иной обстановке. Таким образом сказываются и влияния биотического окружения на организм животного через «упражнение и неупражнение» его органов. В первую очередь здесь идет речь о мышцах, объем которых увеличивается при тренировке (у позвоночных животных), затем о структуре, а частью и о форме костей, меняющихся в зависимости от нагрузки. Кроме того, хотя и в менее выраженной форме, это касается легких, почек, желудка и кишечника, сердца и сосудов, кожных покровов (мозоли) и других органов. Наконец, в более тонкой форме это касается и органов чувств и центральной нервной системы. В этом последнем случае медленно обратимые морфогенетические процессы установления нервных связей сочетаются теснейшим образом с особенно быстро обратимыми физиологическими реакциями.
Обратимость модификаций, хотя и была выдвинута нами для проведения известных граней между морфогенетически-ми реакциями (которые могут касаться тончайших структур, как в последних примерах) и физиологическими (которые всегда связаны и с временными изменениями в тонких структурах), бывает выражена весьма различно. Иногда модификации легко обратимы в пределах одной и той же особи и могут повторяться сколько угодно раз в любом направлении.
Растение той расы китайского первоцвета, которая характеризуется белыми цветами при комнатной температуре и красными—в оранжерее (при 30°), можно перенести из комнаты в оранжерею: ближайшие же цветы будут красными; можно ту же особь перенести обратно в комнату, и следующие по порядку цветы будут белыми.
У животных некоторые изменения окраски отличаются очень быстрой обратимостью. Тогда говорят о физиологических реакциях (окраска камбалы под цвет дна, окраска хамелеона). В других случаях они лишь медленно обратимы (у многих земноводных) или в пределах особи совсем необратимы (зависимая пигментация куколок и взрослых насекомых). Тогда говорят о модификациях. К медленно обратимым реакциям относятся и результаты «упражнения и неупражнения» органов. Результаты тренировки мышц угасают при прекращении тренировки. Структура губчатого вещества кости всегда перестраивается соответственно нагрузке данного времени. Это процесс не быстрый, но он может менять свое направление, т. е. обратим в пределах особи. То же самое касается густоты шерстного покрова млекопитающих, оперения птиц, изменений сердца, легких и кишечника позвоночных. Другие модификации, как уже упомянуто, развившись у известной особи, удерживаются ею затем на всю жизнь.
Обратимость модификаций, и особенно адаптивных модификаций, является их существенным свойством. Если способность к адаптивным модификациям рассматривать как исторически развившееся приспособление к изменчивым факторам внешней среды, а иначе их трудно расценивать, то весь их смысл, вся их ценность как приспособления, заключается именно в их. обратимости. Вне этой обратимости, т. е. ненаследственности, не было бы и самого индивидуального приспособления.
Каждый факт унаследования конкретной модификации, если бы он был возможен, уничтожал бы ее адаптивность для потомков, попадающих в иные условия существования, отличающиеся от родительских по тем факторам, которые определяли развитие этой модификации.
3. НОРМА РЕАКЦИИ
Любой генотип характеризуется своей опреде-леннойнормойреакций. В состав этой нормы входят и индивидуальные реакции организма, попадающего в различные условия внешней среды, т. е. способность к различным его модификациям. В случае резко выраженной целостности адаптивных модификаций организма можно говорить об «адаптивных нормах», как частных проявлениях общей нормы реакций. Такими адаптивными нормами являются «экофены» Турессона как формы индивидуального приспособления растений к свойствам почвы и другим экологическим условиям. Такими же адаптивными нормами будут альпийский габитус многих растений, а также водная и воздушная формы амфибиотических растений, сезонные формы многих животных и т. п.

Рис. 1. Некоторые мутации Drosophila melanogaster, выражающиеся главным образом в нарушении нормального строения крыльев. 1 — Notch, 2 — Beaded, 3 — rudimentary, 4 — curled, 5 — vestigial. Все самки. По Т. Моргану, 1919, и Т. Моргану и др., 1925
Совершенно иной характер имеют неприспособительные модификации, или «морфозы». Они возникают как новые реакции, не имеющие еще своей исторической базы. При этом либо организм сталкивается с новыми факторами среды, с которыми он в своей истории не имел дела, либо, в результате мутации, норма реакций самого организма оказывается измененной (нарушенной). В первом случае мы наблюдаем «рентгеноморфозы», «хе-моморфозы», «термоморфозы» и т. п.. во втором случае — различные выражения мутаций при разных условиях внешней среды. Все эти реакции в лучшем случае индифферентны, чаще всего они имеют характер более или менее резко выраженного нарушения существующей организации или проявляются как явные уродливости, мало жизнеспособные в естественных условиях существования. Такие морфозы, как правило, очень лабильны: они весьма легко меняют свое выражение при изменении интенсивностей факторов внешней среды. Примерами морфозов могут служить изменчивые выражения многих мутаций плодовой мушки-дрозофилы.
Известная мутация уродливого брюшка (Abnormal abdomen) получает свое максимальное выражение при развитии личинок на хорошем влажном корме. По мере его подсыхания выражение меняется вплоть до почти нормального строения брюшка. Мутация vestigial обладает при высоких температурах (32°) почти нормальными крыльями (по крайней мере, у известного числа особей); при понижении температуры их величина уменьшается, и они становятся совершенно рудиментарными при 25–18° С. Аллеломорфная vestigial мутация pennant обладает, наоборот, при низкой температуре почти нормальными крыльями, а при повышении температуры (от 16 до 30° С) они постепенно уменьшаются в размерах. Гетерозигота pennant X vestigial обладает сменой направления реакции. При повышении температуры от 16 до 22° величина крыльев уменьшается, при температурах 22–26° С она остается неизменной (уменьшенной), а при дальнейшем повышении температуры до 32° размеры крыльев возрастают (рис. 22–24). Во всех этих реакциях нет ничего благоприятного, ничего приспособительного. Они в такой же мере нарушают строение организма, как и другие мутации, обладающие более стабильным выражением.
Действительно, новые реакции организма никогда не имеют приспособительного характера. Они возникают вместе с мутациями как различные формы их выражения при разных условиях внешней среды. Если организация растений и животных, строясь на мутациях, приобретает приспособительный характер лишь в процессе эволюции, то и все их реакции, строясь также на мутациях, приобретают свой приспособительный характер также лишь в течение своего исторического развития в определенных условиях внешней среды.

Рис. 2. Рецессивная мутация tetraptera с очень изменчивым выражением. а —нормальный гальтер (жужжальце): b — g — различные выражения мутации; b — е — стадии преобразования гальтера в крыло, f — g — редукция гальтера. По Н. В. и Н. А. Тимофеевым-Ресовским, 1933
Все новое, в том числе и способность к адаптивным модификациям, создается в процессе естественного отбора мутаций с их характерными нормами реакций. Об этом мы еще будем говорить в дальнейшем. Здесь же отметим одно различие между морфозами и адаптивными модификациями. Морфозы, как мы только что видели на ряде примеров, весьма лабильны (как и мутации). Они легко меняют свое выражение при изменении факторов среды. Вместе с тем сходные реакции легко меняют свое выражение и в разных генотипах, даже при небольших различиях (например, Bar и infra-bar, vestigial и pennant обладают противоположными температурными реакциями). Нередко обнаруживаются и значительные индивидуальные различия в выражении мутаций (vestigial, Bar, Abnormal abdomen, Beaded, aristopedia, tetraptera и другие мутации при одних и тех же условиях дают индивидуально различные реакции). В этом ясно проявляется малая устойчивость молодых, еще недавно возникших, форм реагирования (рис. 1, 2).
С другой стороны, конкретные адаптивные модификации иногда весьма устойчивы (как и норма). Они проявляются в общем почти одинаково у самых различных особей данного вида и нередко допускают в довольно широких пределах изменения интенсивностей даже тех факторов среды, которые непосредственно связаны с данной индивидуальной адаптацией. Обычно требуется лишь некоторый минимум интенсивности для того, чтобы известная модификация приобрела свое полное выражение. Дальнейшее повышение интенсивности детерминирующего или освобождающего фактора не приводит к изменению этого выражения. Это ясно указывает на существование внутренних механизмов, регулирующих развитие модификации. В процессе эволюции все формообразование, т. е. весь реакционный аппарат вместе со всеми его приспособительными реакциями, снабжается регулирующими механизмами, защищающими процессы индивидуального развития от возможных нарушений их со стороны изменчивых, и во многом случайных, влияний факторов внешней среды. Элементы авторегуляции характерны для всех адаптивных модификаций и отличают их как процессы, имеющие за собой длительную историю, от морфозов как реакций, не имеющих еше своего исторического прошлого. И адаптивность модификаций, и их устойчивость есть результат эволюции в известных условиях внешней среды.
Понятие «нормы реакций» имеет большое значение для общих проблем биологии, как одно из немногих строго определенных понятий, которые помогают внести полную ясность в дискуссионные вопросы о формах изменчивости и их значении в процессе эволюции. Любая модификация означает определенную форму индивидуального реагирования, входящую в норму реакций данного организма и являющуюся одной из ее характеристик. С другой стороны, мутация означает изменение этой нормы реакций организма, и это является исчерпывающей характеристикой любой как «видимой», так и «физиологической» мутации.
4. МУТАЦИИ И ИХ ВЫРАЖЕНИЯ
Как уже сказано и как это видно из приведенных примеров, мутации выражаются в наследственном изменении нормы реакций. Это есть их наиболее общее и всегда правильное определение.
Изменение нормы реакций означает обычно и изменение процессов индивидуального развития (которые определяются именно нормой морфогенетических реакций), т. е. более или менее глубокое изменение организации, вплоть до тончайших структур, а также любых признаков и функций организма. Наиболее изучены мутации у плодовой мушки-дрозофилы (Drosophila melanogaster). Здесь известно множество мутаций с измененной окраской тела (от почти черной до светло-желтой) или отличным цветом глаз (кроме нормальной красноглазой формы, известны мутации с темно-бурыми, вишневыми, киноварными, эозиновыми, кремовыми и белыми глазами), с изменением длины или формы щетинок, с различным их расположением: некоторые щетинки или целые группы щетинок могут выпасть или появляются удвоенные, или добавочные щетинки на новых местах. В мутациях изменяются величина и форма глаз, строение глазков, величина и форма крыльев (рис. 3), длина и строение антенн и конечностей, строение ротовых частей, груди или сегментов брюшка. Известны безглазые и бескрылые формы. Изменяются части наружного полового аппарата, а также и половые железы с их придаточными частями. Мутации могут различаться по числу яйцевых трубок, по числу яйцевых камер, по внешнему виду самих яиц. Мутации выделяются нередко по пигментации внутренних органов или по их недоразвитию.
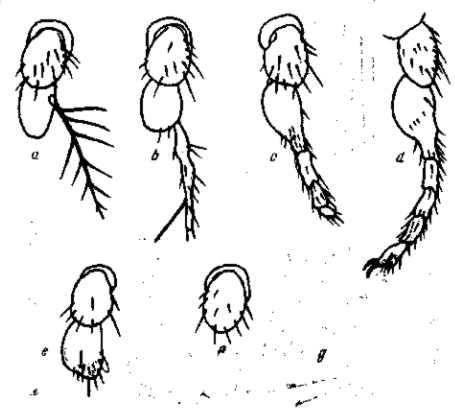
Рис. 3. Рецессивная мутация aristopedia с очень изменчивым выражением. а — нормальная антенная Drosophila melanogaster; b — g — различные выражения мутации: b — d— преобразование антенны в ножку, е — f — редукция антенны, g —полное исчезновение антенны. По Н. В. и Н. А. Тимофеевым-Ресовским, 1933
Головной мозг изменяется в связи с изменением величины глаз и т. п. Иногда вместо гальтеров развивается пара крыльев, вместо антенн — конечности. Мутации отличаются большей или меньшей плодовитостью (числом откладываемых яиц), большей или меньшей длительностью эмбрионального или личиночного развития, по скорости роста личинки и по числу ее линек. Мутации характеризуются и по своей общей «жизнеспособности» и по средней длительности жизни. Эти последние мутации принято называть «физиологическими». Многие мутации связаны с более значительными нарушениями процессов индивидуального развития, очевидно с расстройством системы корреляций, которые приводят к более глубоким ненормальностям, к остановке развития и, рано или поздно, к гибели развивающегося организма. Такие мутации называются «летальными». Иногда при этом нарушенными оказываются уже процессы образования и развития яйца, процессы дробления или гаструляции; в других случаях расстраиваются процессы образования органов, их координация и физиологические корреляции; в результате организм оказывается действительно нежизнеспособным в любых условиях развития и существования.
Как видно, разные мутации характеризуются определенными качественными различиями. Кроме того, они отличаются друг от друга по глубине захвата основ самой организации, по широте охвата различных ее сторон и по степени своего выражения. Мы знаем у дрозофилы множество мутаций, выражающихся в недоразвитии крыльев (см. рис. 1). Различные мутации связаны при этом с самым различным выражением этого недоразвития, начиная от очень небольшого уменьшения размеров крыловой пластинки или образования на крыльях небольших вырезов вплоть до полной рудиментации крыльев, со всеми возможными переходами между наличием нормальных крыльев и их полным отсутствием (см. рис. 15). Мутации могут быть крупными и малыми, глубокими или поверхностными, морфологическими или физиологическими. Иногда они оказываются в своем выражении довольно узко локализованными; в других случаях они распространяются на множество различнейших признаков. Все то, что в таких деталях изучено у дрозофилы, оказывается справедливым и для других животных и растений (из них некоторые, как, например, львиный зев, изучены также с весьма значительной полнотой) . Мутационный процесс протекает по сути одинаково у различных организмов.
Из многочисленных мутаций, которые могут послужить материалом для прогрессивной эволюции, мы должны особо выделить весьма обычные мутации, выражающиеся в увеличении числа сходных образований (полимеризация). Такие изменения создают почву для дальнейшего усложнения организации путем разделения функций и соответственной дифференциации го-мономных частей.
Мы рассмотрим здесь несколько ближе лишь некоторые вопросы, связанные с мутационной изменчивостью и имеющие большое значение для понимания эволюционных процессов:
A. Вопрос о жизненности мутаций и их характеристике как прогрессивных и регрессивных изменений организма. Мутации «жизнеспособности».
Б. Вопрос о плейотропии, т. е. о множественном или многостепенном выражении мутаций.
B. Вопрос об изменчивости их выражения, включая вопрос о непостоянстве проявления и о возможности скрытого мутирования.
Г. Вопрос об источниках мутаций по частоте мутирования. Мутабильность.
А. ЖИЗНЕСПОСОБНОСТЬ МУТАЦИЙ
Огромное большинство мутаций представляют собой более или менее резко выраженные нарушения строения организма и его функций. Это не должно нас удивлять. Все организмы обычно приспособлены к той нормальной обстановке, в которой они живут, и это объясняется их историческим формированием именно в этой обстановке, т. е. в известных, совершенно определенных, условиях внешней среды с ее физическими факторами (почвы, климата и т. п.) и биотическим окружением. Кроме того, любая организация отличается внутренней согласованностью структур и функций. При сложности как внутренних взаимозависимостей частей организма, так и связей его с внешней средой вполне естественно, что почти всякое изменение организации в целом или в ее частях и функциях оказывается неблагоприятным — оно всегда нарушает исторически установившиеся взаимозависимости как в самом организме, так и в его соотношениях с факторами внешней среды. Можно прямо сказать, что при данных условиях внешней среды, к которым данный организм вполне приспособлен, любая мутация оказывается вредной, так как она меняет установившиеся соотношения и, следовательно, нарушает исторически создавшуюся приспособленность организма к этой среде. Само собой разумеется, что масштаб этих нарушений определяется как глубиной мутационного изменения, так и его локализацией. Малые мутации могут быть связаны с очень небольшими нарушениями организации. Их вредность снижается еще более, если мутация охватывает мало ответственные, «поверхностные» признаки и не нарушает заметно более существенных основ организации и жизненно важных приспособлений. Поэтому малые мутации могут и не ставить их обладателей перед особыми опасностями. Такие мутации могут до известной степени размножаться в популяции.
Кроме того, следует обратить особое внимание на то обстоятельство, что факторы внешней среды, к которым приспособлен известный организм, не бывают неизменными (особенно биотические факторы). При изменениях в факторах внешней среды организм теряет свою к ним приспособленность. В этом случае может возникнуть такое положение, что мутация, меняющая те признаки, которые имеют известное отношение к изменившимся факторам среды, может оказаться в более благоприятном положении, чем прежняя, нормальная форма. Мутация, бывшая ранее вредной, может при изменении внешней среды приобрести, положительное значение. Это первое существенное примечание к общему положению о большей или меньшей вредности мутаций.
Вредность мутаций имеет, следовательно, относительное значение. Можно говорить об условной вредности мутаций (в данных условиях существования) и об условной их полезности (при некоторых иных условиях).
Второе, еще более существенное обстоятельство, на которое нам следует обратить внимание, состоит в том, что нормально особи любого вида живут в разнообразных и изменчивых условиях внешней среды. Как физические факторы, так и биологические бывают различными в различных местообитаниях одного и того же вида организмов, а также подвержены как случайным, так и периодическим (в частности, суточным, сезонным) изменениям. Это ставит один и тот же вид организмов в различные условия в разное время и в разных местностях. То, что было вредным в одних условиях, может оказаться безразличным или даже благоприятным в других. В этих случаях мы будем говорить об условной вредности и условной полезности мутаций.
Наконец, в-третьих, при множественном выражении каждой отдельной мутации, может оказаться, что мутация, в общем вредная, обладает все же и безразличными, а иногда, быть может, и отдельными относительно благоприятными выражениями. Эти последние могут в некоторых случаях приобрести положительное значение в известных условиях жизни организма. О таких мутациях можно говорить как о частично полезных и частично вредных.
Вредность мутаций проявляется в их малой жизнеспособности в природных условиях существования. Опыты с выпуском мутан-тных мух-дрозофил на волю показали их быстрое исчезновение (Gordon С, 1935; Гершензон, 1941б).
Генетики иногда говорят о полной «жизнеспособности» тех или иных мутаций. В данном случае имеется, однако, в виду жизнеспособность в условиях экспериментальной культуры, при полной обеспеченности подходящим кормом, при оптимальных физических условиях и при совершенной защите от каких-либо врагов или паразитов. Как бы ни были «жизнеспособны» мухи с рудиментарными или уродливыми крыльями, с кривыми ногами или дефектными глазами, они при жизни на воле в природных условиях быстро погибли бы от голода и от хищников. Кроме того, обычно значительно снижена и плодовитость мутантных мух.
Однако даже в оптимальных условиях лабораторных культур плодовитость и «жизнеспособность» большинства мутаций оказывается более или менее значительно пониженной (не говоря уже о многочисленных совершенно нежизнеспособных «летальных» мутациях).
На прилагаемой таблице 1 сопоставлены данные по жизнеспособности некоторых мутаций Drosophila funebris, выраженные в процентах от жизнеспособности дикого типа (по Тимофееву-Ресовскому) .
ТАБЛИЦА 1. Жизнеспособность некоторых мутаций и их комбинаций у Drosophila funebris (в % от жизнеспособности дикого типа той же мухи)
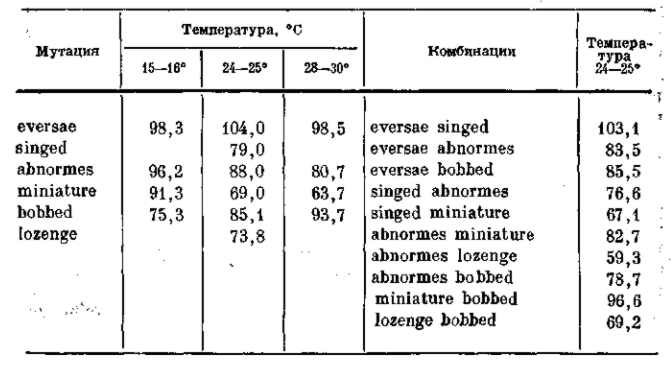
Из этой таблицы видно, что большинство мутаций связано со снижением жизнеспособности в условиях лабораторной культуры. Исключение составляет лишь мутация eversae, которая в этих условиях, при температуре 24–25°, оказывается несколько более жизнеспособной. Однако уже в условиях несколько большей плотности населения их жизнеспособность становится ниже нормы. С другой стороны, мутация bobbed обладает в уплотненных культурах более высокой жизнеспособностью, чем в нормальных. Это является некоторой иллюстрацией к сказанному ранее об условной вредности мутации. Необходимо обратить внимание и на то, что некоторые мутации при их комбинировании обнаруживают резко сниженную жизнеспособность (abnormes lozenge), а другие мутации, которые в отдельности отличаются сниженной жизнеспособностью, как miniature и bobbed, в комбинации могут восстановить почти нормальную жизнеспособность дикой формы. То же самое было показано Кюном (Kuhn, 1935) для некоторых мутаций амбарной огневки Ephestia kiihniella. О значении таких фактов мы будем еще говорить в дальнейшем. Условность вреда мутаций может быть показана и на следующем примере: веслоногий рачок Daphnia longispina обладает температурным оптимумом в 20° (по Banta, Wood, 1927) и при температуре 27° может прожить лишь несколько дней. Авторам удалось установить существование мутации этого рачка, которая гибнет в «нормальных» температурных условиях, т. е. при 20°, но имеет свой температурный оптимум между 25 и 30°.
Б. МНОЖЕСТВЕННОСТЬ ВЫРАЖЕНИЯ МУТАЦИЙ
Хотя наименование определенных мутаций указывает на изменение того или иного признака, обычно внешнего, легко бросающегося в глаза, можно определенно сказать, что мутаций, меняющих только один признак, в действительности не существует. Если морфологическое выражение мутации иногда как бы концентрируется на определенных признаках, оставляя другие признаки малоизмененными, то любая мутация все же всегда имеет и известное физиологическое выражение в виде нарушения некоторых функций, изменения процессов развития и роста, изменения плодовитости или общей жизнеспособности.
Еще Мендель нашел, что у гороха красная окраска цветов связана с развитием красных пятен в пазухах листьев и с серой окраской кожуры семян. По Нильсону-Эле, у овса окраска чешуи и длина ости изменяются совместно с изменением одного и того же наследственного фактора. У китайского первоцвета (Primula sinensis) известна мутация (Ch), у которой лепестки разрезаны, число чашелистиков удвоено, все растение имеет более компактный вид и, в комбинации с некоторыми другими изменениями, несколько изогнутую форму листьев.
У дрозофилы (D. melanogaster) мутация рудиментарных крыльев vestigial характеризуется уменьшенной величиной крыльев; однако, кроме того, она отличается положением щетинок, изменением гальтеров, уменьшенной длительностью жизни, уменьшением числа яйцевых трубочек и пониженной плодовитостью. Сходная мутация rudimentary обладает, кроме рудиментарных крыльев, также укороченными задними ножками. Мутация short-wing отличается различной степенью редукции крыльев, уменьшением и «погрубением» глаз (рис. 4). Мутация укороченных ножек dachs обладает и измененным жилкованием крыльев. Мутация изогнутых крыльев bent имеет и укороченные, согнутые ножки, нарушенное расположение щетинок на груди и отличается «погрубением» глаз. Мутация stubbloid характеризуется укорочением щетинок, но, кроме того, отличается и по строению крыльев, конечностей, антенн и сниженной жизнеспособностью. Мутация белых глаз (white) сопровождается изменением общей окраски тела, укорочением крыльев и изменениями в половом аппарате. У амбарной огневки (Ephestia kuhniella) известна красноглазая мутация, которая отличается, кроме того, измененной пигментацией личинки (светло-зеленая кожа вместо обычной красноватой; более светлые, но все же черные глаза), бесцветными семенниками, замедленной скоростью развития.
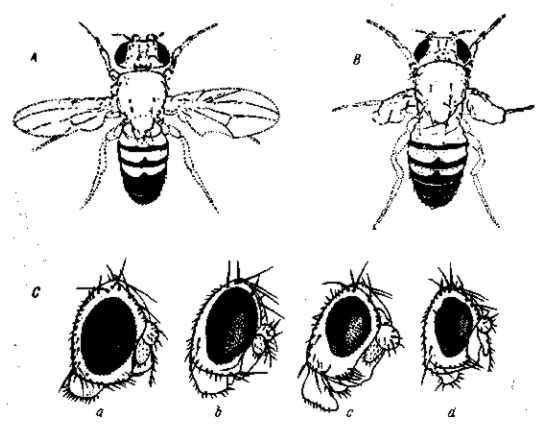
Рис. 4 Drosophila melanogaster. Рецессивная мутация short-wing с плейотропным и очень изменчивым выражением недоразвития крыльев (от нормальных до рудиментарных) и изменения их жилкования, уменьшения величины и погрубения глаз и понижения жизнеспособности. Проявление зависит от температуры: при высоких температурах мутация летальна: при 27,5° типичные ее выражения захватывают 100% всех особей; при понижении температуры процент проявления падает и при 14° все мухи неотличимы от нормальных «диких». А — самец sw с растопыренными, изогнутыми вниз крыльями, характеризующимися, кроме того, вырезками и измененным жилкованием; В — самец с уродливыми рудиментарными крыльями; С — глаза самцов с различным выражением уменьшения и погрубения глаз (a — фенотипически нормальный глаз). По Р. Экеру, 1935
У домашних кур оперение ноги сопровождается изменением чешуи на цевке, укорочением четвертого пальца. Изменение формы гребня, и особенно его раздвоение, сопровождается изменением в строении носовой перегородки, носовых костей и в расположении ноздрей. Появление хохла на голове сопровождается изменением крыши черепа. Курчавость кур связана с пониженной жизнеспособностью. Криперы (коротконогость) характеризуются глубоким нарушением процессов образования кости. В гомозиготном состоянии они нежизнеспособны. Интересно, что здесь, как и во многих других случаях (например, желтые мыши), при рецессивности летального проявления мутации, видимое выражение той же мутации (коротконогость кур, желтая окраска мышей) оказывается доминантным. То же самое установлено для очень многих доминантных мутаций дрозофилы (Beaded, Notch, Dichaete, Star, Truncate и др.) Эти явления плейотропии объясняются существованием коррелятивных связей между различными процессами индивидуального развития. Так как коррелятивные связи охватывают частью внутриклеточные биохимические процессы, которые простираются на различные ткани и зачатки органов и выражаются в их контактном взаимодействии, гормональной обусловленности различных процессов, во взаимном перемещении клеточного материала или в зависимых двигательных реакциях целых пластов и т. п., то и выражение плейотропии может быть весьма разнообразным.
Мы можем выделить две существенно различные формы плейотропии: плейотропию с множественным (полиморфным) выражением и плейотропию с многостепенным (полифазным) выражением.
Множественное выражение плейотропии. Первично, очевидно, любое наследственное изменение должно было изменять весь организм в целом. Если мутация определяется изменением в хроматине ядра, то она должна выражаться прежде всего в изменении течения процессов клеточного обмена, регулируемых взаимодействием ядра и плазмы в каждой клетке тела. В недифференцированном организме с однородными клетками такое изменение, очевидно, сказалось бы одинаково на всех клетках тела, В дифференцированном организме с разнородными клетками процессы внутриклеточного обмена протекают различно в различных тканях и органах. Естественно, что и при изменении процессов обмена в результате мутации это изменение скажется по-разному в различных тканях и органах. Мутация получит множественное выражение. С другой стороны, если разные части организма находятся в разном положении относительно факторов внешней среды, то процессы обмена и в них идут также несколько различно. И в таком случае мутация получит в этих частях различное выражение.
Изменение одного «гена», сказывающееся так или иначе во всех клетках организма, получает различное выражение в зависимости от наличных дифференцировок и различий в положении и от физиологического состояния тех или иных частей организма.
Примерами такого множественного (полиморфного.— Ред.) выражения плейотропии и являются многочисленные, частью приведенные выше (рис. 4), мутации дрозофилы. Локализация изменений объясняется в данном случае именно наличной диффе-ренцировкой клеток в пределах известной ткани или органа.
Многостепенное (полифазное.— Ред.) выражение плейотропии. Плейотропия получает многостепенное выражение в случае наличия более сложных взаимозависимостей между частями развивающегося организма. В этом случае изменение, определяемое наличной уже дифференцировкой, оказывает косвенное влияние на другие, позднейшие дифференцировки, зависящие в своей реализации от первоначальной дифференцировки. Первичное изменение в процессах внутриклеточного обмена оказывает влияние на течение известного морфогенетического процесса. При наличии более или менее сложных коррелятивных зависимостей изменение одного морфогенетического процесса влечет за собой изменение другого, связанного с ним процесса. Это вызывает, в свою очередь, дальнейшее изменение в процессах, зависящих от предшествующих процессов. Таким образом реализуется последовательная цепь изменений, следующих друг за другом и обусловливающих друг друга. Изменение одного «гена», сказывающееся, конечно, во всех клетках организма, получает различное выражение в зависимости от наличных дифференцировок, но, по мере дальнейшего развертывания процессов индивидуального развития, эти различия не только углубляются, но и усложняются вместе с каждым возникновением новых морфологических процессов, зависящих от предшествующих. Мутация получает все более сложное выражение, соответственно степеням усложнения самой организации. Примерами многостепенной плейотропии могут послужить некоторые мутации у позвоночных животных.
Мохноногость кур определяется ранней концентрацией мезенхимы под кожей постаксиального края конечностей. Это первое видимое изменение. В связи с этим стоит нарушение нормального расположения зачатков чешуи на цевке, а затем закладка ряда перьев. Это — вторичные видимые изменения процессов развития. Мезенхима, идущая на образование закладок перьев, имеет, однако, скелетогенное происхождение, она извлекается из закладки постаксиальной части скелета конечности, и, в связи с нехваткой этого материала, более или менее недоразвиваются фаланги 4-го пальца (брахидактилия), палец изгибается на тыльную сторону и на нем недоразвивается коготь (рис. 35). Об этих изменениях можно говорить как о третичных выражениях мутации мохноногости. Все эти изменения представляют ступени последовательного усложнения выражения одной простой мутации.
Но этот плейотропизм имеет явно иной характер, чем то, что мы знаем у дрозофилы. Там мы говорим о множественном выражении мутации, понимая под этим только различную локализацию отдельных изменений. Здесь мы говорим о многостепенном выражении, поскольку локальное первичное изменение влечет за собой ряд дальнейших изменений возрастающей сложности.
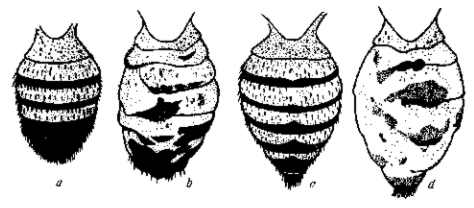
Рис. 5. Доминантная мутация уродливого брюшка Abnormal abdomen у Drosophila melanogaster. Выражение этой мутации зависит от влажности питательной среды, в которой выращиваются личинки. а, b — нормальное и мутантное брюшко самца; с, d — то же у самки. У гетерозиготы выражение мутации теряется (т. е. брюшко становится нормальным) при подсыхании корма. Доминантная мутация превращается тогда в рецессивную. По Т. Моргану, 1919
В. ИЗМЕНЧИВОСТЬ ВЫРАЖЕНИЯ МУТАЦИЙ
Выражение мутаций в той или иной мере изменяется как в зависимости от индивидуальных особенностей организма (его генотипа), так и в зависимости от факторов внешней среды. Эта изменчивость может иметь до известной степени качественный характер или касается, главным образом, количественной стороны выражения известного признака (рис. 3, 4, 5). В этих случаях говорят о степени выражения или экспрессивности мутации. Однако эта степень выражения может снижаться до нуля, так что несомненно имеющаяся мутация у данной особи ни в чем не выражается. Мутация может находиться в скрытом виде у многих особей. В группе однородных по происхождению мутантов сама мутация получает заметное выражение лишь на отдельных особях, составляющих известный процент всех особей. В таком случае говорят о проценте проявления или пенетрантности мутации.
Выражение мутации изменяется с изменением факторов внешней среды. Это ясно видно на многих мутациях дрозофилы. Мы уже упоминали о мутации уродливого брюшка (Abnormal abdomen), которая получает свое типичное выражение лишь при воспитании личинок на свежем корме (рис. 5), о мутациях лентовидных глаз Ваг (рис. 6, 7) и о мутациях недоразвитых крыльев vestigial и pennant, меняющие выражение при разных температурах. Мутация eyeless меняется в зависимости от качества корма. Меняется и физиологическое выражение мутаций — плодовитость и жизнеспособность при разных температурах и т. п. В особенности изменчиво обыкновенно выражение мутаций в гетерозиготе. Даже вполне рецессивные мутации могут получить заметное выражение, например, при повышенной температуре (Child, Blanc, Plough, 1940). Выражение мутации меняется и в зависимости от гормональных влияний. Это видно на мутациях, касающихся зависимых вторичнополовых признаков у птиц (кур) и у млекопитающих. Так, например, комолость получает более резкое выражение у овец, чем у баранов. В особенности это опять-таки касается мутаций в гетерозиготном состоянии.
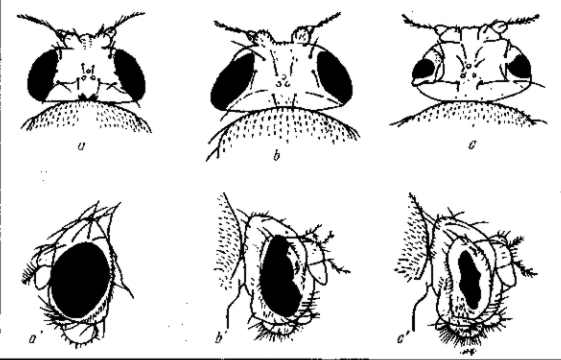
Р и с. 6. Доминантная мутация полосковидных глаз Ваг у Drosophila melanogaster. Выражение этой мутации сильно меняется в зависимости от температуры, при которой воспитывались личинки. а, а' — нормальные глаза; с, с' — полосковидные глаза; b, b' —гетерозиготная мутация. По Т. Моргану, 1919
Однако выражение мутации меняется и в зависимости от наследственных свойств организма, т. е. оно оказывается разным в различных генотипах. Форд и Гексли (Е. Ford a. L. Huxley, 1927) показали, например, что у бокоплава Gammarus chevreuxi одна и та же мутация выражается в одной линии в появлении красных глаз вместо черных, а в другой линии — в появлении коричневых глаз. У дрозофилы мутация beadex обладает в гетерозиготном состоянии различным выражением в разных генотипах (Д. М. Шифрин). Мёллером и другими исследователями было уже давно установлено для целого ряда мутаций дрозофилы (D. melanogaster), отличающихся сниженной жизнеспособностью, что при длительном их культивировании в лабораторных условиях морфологическое выражение мутаций постепенно угасает, а вместе с тем восстанавливается и нормальная жизнеспособность мух. Такова, например, мутация безглазия (eyeless, рис. 21), которая в чистых культурах довольно быстро теряет свое выражение, причем восстанавливаются нормальное строение глаз и нормальная жизнеспособность. Мутация переходит в скрытое состояние, но не исчезает, так как при скрещивании с другой такой же линией (eyeless, потерявших свое выражение) она вновь проявляется. Мутация stubbloid характеризуется укорочением щетинок, сморщенными крыльями, изогнутыми ножками и сниженной жизнеспособностью. В чистых (инбредных) культурах этой мутации изменение крыльев и изгиб ножек со временем становятся все менее заметными и в то же время общая жизнеспособность мух постепенно восстанавливается (Добжанский, 1937). Скрещивание с нормальными «дикими» мухами ведет к расщеплению и восстановлению характерного выражения мутации (во втором поколении). Некоторые мутации оказываются особенно неустойчивыми в своем выражении: они очень чувствительны как по отношению к внешним факторам, так и по отношению к индивидуальным свойствам генотипа. Такие мутации характеризуются обычно по среднему их выражению. Так, например, мутация лентовидных глаз Ваг может быть охарактеризована лишь по общей форме глаза и по среднему числу фасеток, так как это число сильно варьирует. Еще более варьирует число фасеток у мутации безглазия (eyeless).
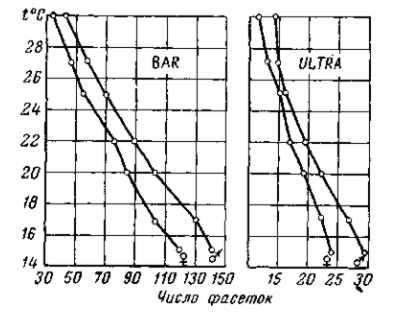
Рис.7. Обратная зависимость величины глаза и числа его фасеток от температуры У мутаций полосковидных глаз Ваг и Ultra-Bar. При повышении температуры от 15 до 30° среднее число фасеток глаза Ваг падает от 120 — 140 до 35 — 40, а среднее число фасеток глаза Ultra-Bar падает от 25 — 30 до 12–15. При этом мутация всегда сильнее выражена у самок, соответственно имеющейся паре Х-хромосом, в которых локализовано данное изменение. У самцов, обладающих лишь одной Х-хромосомой, число фасеток глаза несколько выше Слева — температурная зависимость выражения Ваг; справа — то же для Ultra-Bar. По Е. Драйверу, 1931
И здесь следует подчеркнуть, что в гетерозиготном состоянии выражение полудоминантных мутаций особенно малоустойчиво. Сдвиги в степени доминирования наблюдаются очень часто как под влиянием факторов внешней среды, так и под влиянием гормональных воздействий, а также в зависимости от данного генотипа.
Малые мутации. Некоторые мутации имеют ничтожное выражение. Существование малых мутаций, обнаруживаемых обычно лишь по их совместному выражению с некоторыми другими мутациями, т. е. по их «модифицирующему» влиянию, было уже давно замечено некоторыми исследователями. Однако лишь относительно недавно на дрозофиле удалось показать, что такие мутации по существу ничем не отличаются от других генных мутаций и основываются на таких же локальных изменениях отдельных хромо-меров («генов» Моргана).
Бриджес (1919) нашел восемь разных наследственных факторов, меняющих окраску глаз у мутации «эозиновых» (eosin) глаз дрозофилы от темно-эозиновых до белых (по виду не отличимых от мутации белоглазия, white). Удалось определить и локализацию этих генов-модификаторов в хромосомах. Благодаря существованию малых мутаций, меняющих выражение других, более значительных, практически почти любая мутация реализуется в виде целого ряда градаций, которые могут быть предметом естественного отбора. Опыты применения искусственного отбора для повышения и понижения выражения определенных мутаций были проведены сначала Стертевантом (1918) у мутации Dichaete дрозофилы по числу щетинок, а затем, с неменьшим успехом, и на многих других мутациях, на различных иных объектах.
Это показывает, что и в природных условиях малые мутации, служащие источниками небольших градуальных различий отдельных особей, должны играть большую роль как основной материал для естественного отбора и эволюции организмов.
Неполное проявление мутаций. Если выражение мутации колеблется между развитием известного признака и его полным отсутствием, то мы имеем перед собою мутацию с неполным проявлением. Степень выражения мутации в известных условиях и в известных линиях учитывают тогда отношением числа особей с развитием характерного признака к общему числу особей данной линии (в данных условиях), выраженным в процентах. Это и будет процент проявления мутации. Хорошим примером плейотропной мутации с весьма изменчивым выражением и неполным проявлением может послужить мутация polymorpha Drosophila junebris. Эта мутация выражается в различной степени недоразвития крыльев (вплоть до рудиментации), в погрубении и уменьшении величины глаз (до почти полной редукции), в уменьшении числа члеников лапки (tarsus) и других изменениях. Проявление всех этих изменений неполное и «резко» меняется в зависимости от качества корма и температуры. При 25° процент проявления достигает по основному выражению 90–100%, т. е. почти все особи оказываются с дефектными крыльями, 70–80% особей имеют при этом уменьшенные и «грубые глаза» и 20–30% особей — укороченные ножки. При 15° процент проявления падает по крыльям до 20–30%, а по глазам и ножкам — до 0, т. е. глаза и ножки всех особей оказываются нормальными. Проявление мутации зависит и от генотипа в целом и от отдельных модификаторов: путем искусственного отбора проявление мутации может быть значительно изменено (Промптов, 1929).
Факт существования мутаций с неполным проявлением показывает, что в некоторых случаях сказывается особенно резко зависимость выражения известного признака от внешних или внутренних факторов развития. В этом случае факторы, детерминирующие известную морфогенетическую реакцию, встречают, очевидно, реагирующую ткань на грани порогового уровня ее раздражимости, и самый незначительный сдвиг в интенсивности или во времени появления морфогенного раздражителя делает реакцию невозможной. Здесь проявляется значение «чувствительной фазы» в процессах развития, когда малейшее нарушение приводит к значительным результатам.
Изменчивость плейотропных выражений мутаций. Если мутация обладает множественным выражением, то обычно все эти выражения оказываются зависимыми от известных внешних и внутренних факторов. Все они могут иногда переходить в скрытое состояние, как мы на это указывали в мутации stubbloid.
Однако некоторые выражения мутации могут оказаться более лабильными и легче подвергаются изменяющему влиянию внешних или внутренних факторов, чем другие. В таком случае можно ожидать даже возможности изменения разных выражений в различном направлении. Во всяком случае, возможны погашение и переход в скрытое состояние только некоторых (в частности — вредных) выражений мутации, в то время как другие (более благоприятные) выражения сохраняются или даже стабилизируются (см. III; 1). в особенности лабильны вторичные и третичные выражения многостепенного плейотропизма. Мохноногость кур бывает и в своем основном выражении (оперении цевки) весьма различно развита. Но в особенности изменчивы вторичные выражения этой мутации: брахидактилия, синдактилия, изгиб четвертого пальца и редукция когтя на нем. Последнее совпадает обычно с максимальным выражением оперения. Однако брахидактилия, синдактилия, изгиб пальца изменчивы в гораздо большей мере, чем сама мохноногость. Это, очевидно, обусловило возможность выведения породы мохноногих кур (фаверолей) с совершенно нормальными пальцами (рис. 34). Эти факты имеют также большое значение для понимания элементарных процессов, лежащих в основе эволюции.
Г. ИСТОЧНИКИ МУТАЦИЙ И ЧАСТОТА МУТИРОВАНИЯ
Мутации изредка возникают в культурах без каких-либо видимых причин только на отдельных особях. Это показывает, что такие отдельные особи либо попали под случайное влияние какого-то редкого агента, либо отличаются особой чувствительностью по отношению к каким-то более обычным влияниям. Если правильно первое предположение, то можно было бы думать о радиоактивном распаде атомов как об источнике мутационной изменчивости. Если правильно второе предположение, то источником мутационной изменчивости могут быть многие факторы внешней среды и, в особенности, необычные их интенсивности или резкие колебания этих интенсивностей (крайние температуры, условия интенсивной инсоляции, крайние пределы влажности, химические влияния солей или не совсем обычных органических соединений, попавших в состав пищи, и т. п.). Экспериментальные данные говорят в пользу обоих предположений, и нужно думать, что конкретные причины, вызывающие мутации, могут быть весьма разнообразными, однако всегда преломляются через индивидуальные свойства данного организма (его генотипа).
С тех пор как Мёллеру (1927) удалось впервые получить экспериментальные мутации путем воздействия рентгеновских лучей на плодовую мушку (D. melanogaster), этот метод получил быстрое распространение и применяется в настоящее время в любой генетической лаборатории для быстрого получения большого числа мутаций. Такое действие оказывает вообще коротковолновая радиация, начиная с ультрафиолетовых лучей. Действием рентгеновских лучей получаются, однако, не какие-либо специфические мутации, а только те самые мутации, которые изредка возникают и «спонтанно», т. е. без видимых причин. Очевидно, специфика результата определяется в основном самим организмом: некоторые изменения встречаются очень редко, другие — гораздо чаще. По-видимому, в наследственном веществе организма имеются более слабые, легко уязвимые и более прочные, сравнительно устойчивые участки.
В этом выражается одно из наследственных свойств организма. Поэтому для данного вида организмов характерны не только совершенно определенные мутации, но и определенные частоты возникновения этих мутаций. Воздействием рентгеновских лучей удается увеличить частоту мутирования дрозофилы в 200 раз, однако соотношение между обычными и редкими мутациями сохраняется в общем неизменным. Поэтому обыкновенно .говорят, что рентгеновские лучи «ускоряют» процесс мутирования. То же самое было найдено и у растений.
Подобное «ускоряющее» действие было затем установлено и для повышенных температур (Мёллер, 1928; Гольдшмидт, 1938), однако в гораздо более слабой степени. Другими авторами были получены экспериментальные мутации под действием иода, аммиака и ряда других химических веществ как у дрозофилы, так и у растительных объектов.
Частота мутирования. На первых этапах развития генетики мутации считались большой редкостью. Казалось, что мутации встречаются чаще лишь у некоторых особо мутабильных объектов (ослинник Oenothera lamarkiana) или, во всяком случае, лишь в течение некоторых особо чувствительных периодов жизни определенного вида организмов. Позднейшие исследования как над растительными (львиный зев, кукуруза и мн. др.), так и над животными объектами (мелкие грызуны, куры, карпы, многие насекомые, но в особенности наиболее в этом отношении изученная плодовая мушка-дрозофила) показали неправильность этого впечатления. Хотя различные организмы обладают весьма различной мутабильностью, отдельные мутации вовсе не являются редкостью. В особенности это выяснилось, когда стали обращать внимание на самые незначительные изменения и когда были разработаны более тонкие методы учета как имеющихся уже в скрытом виде, так и вновь возникающих мутаций.
Трудности учета всех мутаций обусловлены целым рядом обстоятельств. Прежде всего определенная мутация, возникая в половой клетке, охватывает всегда лишь некоторую частицу одной хромосомы. Поэтому она после оплодотворения оказывается в гетерозиготном состоянии. Когда мутация возникает в диплоидной клетке (в гониях, в зиготе или соматической клетке), она также простирается лишь на элементы одной хромосомы, оставляя гомологичную хромосому нетронутой. Поэтому новые мутации всегда гетерозиготны и в случае их рецессивности не имеют заметного выражения. Кроме этого, в организме имеются и другие регуляторные механизмы, благодаря которым небольшие мутации могут совершенно не проявляться. Наконец, и более крупные доминантные мутации и даже мутации в гомозиготном состоянии могут, переходя из поколения в поколение, потерять свое выражение через процесс комбинирования с другими мутациями. Таким образом, в любой популяции однородных как будто особей скрывается много различных мутаций, присутствие которых может быть обнаружено лишь путем сложного генетического анализа.
Применение такого анализа действительно позволило вскрыть в природных популяциях множество мутаций с более или менее погашенным выражением и позволило учитывать частоту возникновения новых мутаций в условиях экспериментальных культур.
Наиболее точные методы учета мутаций выработаны для дрозофилы (Мёллером, а затем и рядом других исследователей). Сравнительно проста методика учета летальных мутаций в половой хромосоме. Самки, обладающие летальной мутацией в половой хромосоме, дают при скрещивании с нормальным самцом потомство с соотношением полов, равным 2 : 1 в пользу самок. Так как самец получает свою единственную Х-хромосому только от матери, то в случае гетерозиготной мутации половина самцов получит нормальную хромосому, а вторая половина — хромосому с деталью, — эти последние самцы не жизнеспособны. Поэтому число самцов оказывается вдвое меньшим, чем число самок. Если же подопытные самки обладают одной Х-хромосомой с деталью, то возникновение новой летали во второй Х-хромосоме приведет к тому, что в потомстве самцов совсем не будет. Точность экспериментов повышается, если Х-хромосома с деталью обладает еще удобно распознаваемой «меткой» (лентовидные глаза — Ваг) и «запирателем», препятствующим перекресту хромосом и, следовательно, обмену их частями.
Этот метод очень удобен для сравнения частоты возникновения мутаций в различных линиях, а также для изучения влияния внешних факторов на частоту мутирования.
Частота возникновения деталей в Х-хромосоме оказывается действительно весьма различной в разных линиях. Линии дикой дрозофилы, взятой из различных местностей, давали в потомстве (одного поколения) от 0,05 до 1,7% деталей. При более высоких температурах культивирования частота возникновения деталей заметно возрастает (по меньшей мере в два раза при повышении температуры на 10°). Под влиянием высоких доз лучей рентгена частота мутирования, как уже упомянуто, может быть повышена в 200 раз.
Методы учета деталей в других хромосомах несколько сложнее. Они дали величины того же порядка. Максимальные частоты несколько выше, достигая, например, 4% особей для второй хромосомы (данные Оленова и Хармац, для Уманской популяции дрозофилы, 1938).
Анализ процессов возникновения видимых мутаций дал величины того же порядка. В половой хромосоме находили в разных линиях (из различных популяций) от 0,07 до 0,23% новых мутаций на одно поколение (данные P. JI. Берг, 1942).
Для нас имеют особый интерес именно «видимые» мутации. При учете мутирования во всех хромосомах общее число новых мутаций должно охватывать в каждом поколении минимум от 0,4 до 1% всех особей. Баур находил у львиного зева в каждом поколении до 10% малых мутаций. Учет малых мутаций у дрозофилы даст, вероятно, подобные же величины. Это означает, однако, при незначительной элиминации малых мутаций, очень интенсивный процесс их накопления в процессе размножения. Природные популяции должны быть буквально насыщены такими мутациями. Анализ генетического состава популяций подтверждает этот вывод.
Для разбираемых нами проблем, касающихся возможных темпов эволюции в одном определенном направлении, такие величины, характеризующие суммарную мутабильность организма, недостаточно показательны. Гораздо большее значение имеет частота повторения одних и тех же мутаций, которые могут получить положительное значение в процессе эволюции.
Как и надо было думать, эти частоты, т. е. темп мутирования отдельной наследственной единицы, весьма различны для различных мутаций (и, кроме того, меняются в соответствии с общим темпом мутирования в различных популяциях). Некоторые мутации возникают чрезвычайно редко, другие появляются довольно часто. Одна из обычных мутаций — «желтое тело» (yellow) возникает в различных популяциях дрозофилы у 0,05% (популяция Дилижана), у 0,08% (Кашира), у 0,13% (Никитский сад) и у 0,18% самцов (Умань; данные Р. Л. Берг). У кукурузы (по данным Stadler) различные мутации, меняющие цвет и состав эндосперма, возникают с частотой от 1 (и менее) на миллион гамет до 500 на миллион (т. е. 0,05%). Это величины того же порядка, что у дрозофилы.
Для обычных мутаций можно в среднем принять вероятную частоту их возникновения примерно в 1 на 100 000 особей (т. е. 0,001%). В действительности, эта частота иногда бывает значительно выше, а для редких мутаций, конечно, гораздо ниже.
Мутации обратимы. Это означает, что в культуре известной мутации может возникнуть новая мутация, возвращающая особь к исходной неизмененной форме. Как правило, обратные мутации (к дикому типу) встречаются гораздо реже, чем прямые уклонения от дикого типа. Однако в некоторых случаях (мутация forked) обратное мутирование к норме идет примерно с той же частотой, как и возникновение мутации от нормальной формы.
Некоторые «гены» обнаруживают особенную неустойчивость и являются источниками весьма многочисленных как генеративных, так и соматических мутаций. В первом случае получается, в результате многих прямых и обратных мутаций, картина полной неопределенности наследования. Во втором случае соматическое мутирование приводит к образованию секториальных и даже мозаичных химер. В этих случаях говорят об особо «лабильном» состоянии «генов». Иногда обнаруживается связь между такой высокой степенью мутабильности и наличием хромосомных транслокаций, однако, по-видимому, это не составляет общего правила. Нередко мутабильность выражается в легком возникновении целого ряда аллеломорфов. Кроме того, сама мутабильность сильно зависит как от данного генотипа, так и от внешних факторов. Все это дает обширное поле для эффективного действия естественного отбора. Хотя эти явления пока еще изучены не в достаточной мере, вряд ли мы ошибемся, если скажем, что они представляют лишь крайнюю степень общей способности «генов» и генотипа к наследственным изменениям.
Предполагается, что, поскольку мутации возникают спонтанно и притом повторно, главным образом в одном направлении (от дикого типа), процесс мутирования означает переход наследственной единицы в более устойчивое состояние. Естественно, что обратный переход — в менее устойчивое состояние — совершается гораздо реже. В таком случае нормальный («дикий») тип организации должен постепенно разрушиться вследствие перехода всех генов в более устойчивое, т. е. мутантное состояние, если бы не было механизма, противодействующего этому процессу. Этот механизм и есть естественный отбор, устраняющий или нейтрализующий огромное большинство всех мутаций. Нормальная организация постоянно разрушается вследствие процесса мутирования, но она же непрерывно воссоздается (на все более новой наследственной основе) в процессе естественного отбора наиболее жизненных особей.
Мутирование есть непрерывно действующий процесс, который несет ответственность за разрушение всего старого, уже отслужившего свой век, но вместе с тем доставляет и материал для построения все новых форм организации.
Наличие распространенного параллелизма мутаций и модификаций (фенокопий) показывает, что и те и другие идут в направлении изменения некоторых особенно мало устойчивых (или мало «защищенных») процессов. Имеются более «слабые места» механизма индивидуального развития, которые легче всего нарушаются как при изменении внешних, так и при изменении внутренних (наследственных) факторов.
Как модификации, так и мутации обнаруживаются через сходные, по-видимому, в своих истоках очень простые, изменения одних и тех же морфогенетических процессов. Различие между ними лишь в том, что исходное и притом, очевидно, мало специфическое влияние заключается в первом случае в факторах внешней среды, а во втором случае — в наследственных факторах.
5. РОЛЬ ВНУТРЕННИХ И ВНЕШНИХ ФАКТОРОВ ПРИ РЕАЛИЗАЦИИ УНАСЛЕДОВАННЫХ ФОРМ И ИХ ИЗМЕНЕНИЙ
А. РОЛЬ ЯДРА И ПЛАЗМЫ
Роль ядра и плазмы в реализации унаследованных форм и их изменений можно установить лишь при сопоставлении эмпирических данных эмбриологии (механики развития и феногенети-ки), генетики и цитологии. Ряд совершенно прочно установленных данных выявляет яркую картину непрерывного взаимодействия ядра и плазмы во всех процессах развития в вместе с тем выясняет основы дифференцировки клетки с ее распределением функций между ядерными структурами и плазмой.
Совершенно ясна ответственная роль хроматинных структур ядра в возникновении и передаче наследственных изменений от родителей к потомству. Эти изменения покоятся на общих изменениях числа хромосом или на строго локализованных изменениях их грубой или тонкой структуры. Они передаются по наследству вместе с их материальными носителями — хромосомами — и являются относительно стойкими. Экспериментальные мутации получаются лишь при применении исключительно сильных раздражителей (лучи рентгена и сублетальные температуры или дозы химических веществ), да и то лишь на незначительном числе особей. Получаемые тогда мутации отличаются еще большей наследственной стойкостью, чем норма, давшая начало этой мутации. Если нормальное состояние гена представить себе как состояние до известной степени устойчивое, то мутация находится в генетически еще более устойчивом состоянии (мутация, обратная к норме, происходит обычно во много раз реже, чем прямая мутация; только в этом смысле мы говорим о генетической устойчивости мутаций; в отношении формообразования организма и его мутабильности мутация всегда менее устойчива, чем норма). Если имеется ряд аллеломорфов, то он означает ряд скачкообразных ступеней — изменений между нормой и крайней мутацией. Наименее устойчивое состояние гена соответствует норме. Если норма поддерживается при менее устойчивых состояниях элементарных единиц ядерной субстанции, то должны быть силы, направленные на создание и сохранение этого положения. В индивидуальном развитии организма такой силой является непрерывное взаимодействие ядра с плазмой клетки, а также система корреляционных зависимостей между тканями и частями организма. В историческом развитии организма такой силой является, очевидно, естественный отбор (определяемый взаимодействием организма и внешней среды), непрерывно действующий против разрушающих тенденций мутационного процесса. В результате менее устойчивое состояние ядерной субстанции соот- . ветствует наиболее устойчивому положению всей организации ее в целом при данных условиях внешней среды.
Эксперименты, проведенные на ранних стадиях развития самых различных животных, ясно показали, что ядерная субстанция распределяется равномерно между бластомерами, а также между позднейшими продуктами клеточных делений. Ядра всех клеток зародыша равнозначны и потому не могут быть ответственны за процессы дифференцировки. Последние покоятся на различиях в плазме, получаемой бластомерами из тех или иных частей яйца (анимальные и вегетативные плазмы, специальные субстанции отдельных областей яйца), а также на различиях, развивающихся на базе исходных количественных различий (градиенты) и вследствие различий в положении бластомеров и клеток в организме и по отношению к внешней среде. Все эти процессы детерминации и дифференцировки и протекают в основном в протоплазме клеток зародыша (ядро они захватывают редко — в наиболее специализированных тканях). В плазме же протекают и все регуляторные процессы (перестройки структуры изолированных бластомеров или зачатков, процессы дифференцировки и передифференцировки). Можно было бы предположить, что весь онтогенез определяется только плазмой. Но это неверно.
Мы уже видели, что специфическая структура плазмы яйца определяется хроматином его ядра еще в период образования яйца в яичнике (правые и левые яйца моллюсков, яйца тутового шелкопряда и др.). Отсюда основы организации яйца, а следовательно и организма, создаются все же при участии ядра. Это касается, однако, не только основ организации, но и деталей. При межвидовой гибридизации морских ежей и других животных наблюдали, что развитие начинается по материнскому типу, т. е. определяется плазмой яйца (организация которой, как мы знаем, создается при взаимодействии с его ядром, соответственно генотипу матери). Однако, начиная, примерно, со стадии гаструляции, постепенно начинает сказываться и отцовское влияние. Это явно указывает на значение плазмы яйца, запасы которой распределяются в процессе дробления и близки к исчерпанию на стадиях гаструляции, когда начинается энергичный синтез протоплазмы уже под влиянием ядер зиготы, т. е. обоих родителей (в это время соответственно и начинается заметный рост зародыша). В случае больших межвидовых различий развитие нередко доходит все же до гаструляции, но на этом и останавливается. Очевидно, процессы синтеза оказались невозможными вследствие биохимической несовместимости отцовского ядра и материнской плазмы.
Опыты Геммерлинга над регенерацией одноядерной водоросли Acetabularia, характеризующейся своеобразной формой зонтика на длинной ножке, показывают, что формообразование зонтика зависит от веществ, имеющихся в плазме стебелька (даже если ядро, помещающееся в основании стебелька у ризоидов, удалено). Однако, как показывают опыты пересадки стебля Aceiabularia mediterranea на содержащее ядро основание A. wettsteini, эти плазматические вещества вырабатываются при участии ядра и получают от него свою видовую специфику — формообразование зонтика идет по типу A. wettsteini, т. е. определяется ядром (зонтик имеет у этих видов различную форму).
Мы можем вывести заключение, что индивидуальное формообразование идет вообще при взаимодействии ядра и плазмы.
Однако проверим, как обстоит дело с теми изменениями нормального формообразования, которые мы называем мутациями.
Что в реализации мутаций руководящее значение имеет именно ядро, видно по фактам соматических мутаций: первое изменение возникает, во всяком случае, в ядре, и лишь под его влиянием изменяются плазма и ее дифференцировки. С мутацией меняется вообще норма реакций организма. Соответственно меняются не только его «нормальное» формообразование, но и индивидуальные модификации этого формообразования, зависящие от изменений факторов среды. За все это ответственно ядро, однако реализация всех этих изменений протекает в плазе при участии формообразовательных веществ, синтезируемых в ней при постоянном взаимодействии ядра и плазмы.
Только представление о клетке как целом позволяет нам выйти из круга кажущихся противоречий.
Если и в передаче по наследству, и в индивидуальном развитии, и в его отклонениях от нормы, лежащих в основе исторического развития организма, клетка выступает как целое, то все же она — дифференцированное целое, и поэтому естественно думать, что в различных процессах руководящая роль может принадлежать той или иной составной части клетки. В действительности, при ближайшем рассмотрении можно установить совершенно определенные различия в значении ядерных структур и плазмы как в онтогенетических, так и филогенетических изменениях.
Плазма является совершенно специфическим субстратом онтогенеза, в котором разыгрываются все взаимодействия, ведущие к детерминации и дифференцированию. В ней же осуществляются и все дифференцировки, зависящие от факторов внешней среды, включая и их варианты — модификации. Во всех этих процессах есть нечто общее — в исторической перспективе они всегда обратимы. Они ограничиваются в своей конкретной реализации всегда фенотипом одной особи. Даже «длительные» модификации имеют самое кратковременное существование. Они быстро исчезают в ряде поколений. И в этой передаче «длительных» модификаций по наследству обнаруживается слитность наследования — их растворение в процессах скрещивания. Модификации по своей природе непрерывны и лишь в процессе эволюции (в выработке целостных адаптивных норм и механизма авторегуляции) могут приобрести скачкообразный характер ограниченных «типичных» реакций.
Плазма, несомненно, очень устойчива — она лишь медленно меняется в процессе эволюции, медленнее, чем ядро. Это видно из результатов межрасовой и межвидовой гибридизации. Если скрещивание между расами или близкими видами невозможно и не дает плодовитого потомства, то это почти всегда результат несовместимости их ядерных структур (см. далее II, 8, В). Несовместимость ядра и плазмы сказывается обычно лишь при более отдаленных скрещиваниях (остановка развития зародыша, невозможность дробления яйца или даже гибель ядра сперматозоида в плазме яйца).
Если говорить о клетке как о системе, то мы должны признать, что она находится в состоянии весьма подвижного, но и весьма устойчивого равновесия. Подвижность и устойчивость этой системы определяются в значительной мере свойствами самой плазмы. О подвижности говорит обратимость процессов индивидуального развития, идущих под руково дящим влиянием плазмы, обратимость модификаций, регулятор-ный характер многих реакций и взаимозависимость (корреляций). Об устойчивости говорит как регуляторный характер многих реакций и взаимозависимостей и обратимость модификаций, так и видовая их устойчивость и медленность исторических преобразований плазмы. О том же, наконец, говорит и слитность модифика-ционных изменений в их наследовании — существование скользящих переходов между этими изменениями, явно связанными с изменениями в плазме (впрочем, изменения типа морфозов — фено-копий — могут иметь и до известной степени дискретный характер, см. Рапопорт, 1941).
Б. ЗНАЧЕНИЕ ВНУТРЕННИХ ФАКТОРОВ ОНТОГЕНЕЗА
Одними процессами взаимодействия ядра и плазмы нельзя объяснить всего онтогенеза. При «мозаичном» развитии доминирующая роль переходит как будто к самодифференцированию клеточного материала, содержащего в себе все основные факторы формообразования. Однако при этом всегда сказывается и взаимное влияние частей организма и влияние внешних факторов на это формообразование. В других случаях к этим взаимодействиям частей переходит руководящая роль в индивидуальном формообразовании («регуляционное развитие»), и тогда в процессах непосредственного взаимодействия ядра и плазмы решается лишь вопрос специфических структур клеточной дифференцировки (реализация уже детерминированных структур идет и здесь путем «самодифференцирования»).
Взаимодействие частей развивающегося организма осуществляется в основном: 1) в системах градиентов, охватывающих гомогенные (эквипотенциальные) плазматические (в яйце) или многоклеточные (в зачатках) структуры, обладающие лишь количественными различиями, нарастающими в определенных направлениях (градиента), и 2) в морфогенетических (в частности индукционных) системах, объединяющих гетерогенные многоклеточные структуры, которые обладают ясными качественными различиями в происхождении, строении и обмене веществ. Мы не можем здесь разбирать все достижения механики развития (см. «Организм как целое»). Отметим, однако, следующее. Если в градиент-системах и в индукционных системах решается вопрос о качестве, времени, месте и объеме известных зачатков и этим самым устанавливаются общие основы организации, то видовая специфика клеточных и тканевых дифференцировок определяется, по-видимому, всегда свойствами самого клеточного материала, т. е. решается в процессе «самодифференцироваиия» при постоянном взаимодействии ядра и плазмы. Это видно из многочисленных опытов гетерогенной трансплантации частей и зачатков зародышей. При этом ясно обнаруживается формообразовательное влияние одних частей на развитие других. При пересадке первичного индуктора (верхней губы бластопора или крыши первичной кишки) от зародыша одного вида тритона на другой (от Triton cristatus на Т. taeniatus, или наоборот) у хозяина на месте пересадки развивается добавочная центральная нервная система. Она развивается под влиянием трансплантата (взятого, скажем, от Т. cristatus). Клетки хозяина (Т. taeniatus) дифференцируются под влиянием чуждых им формообразовательных веществ (от Т. cristatus). Однако добавочная нервная система, развиваясь на ненормальном месте под чуждым влиянием, получает все специфические особенности структуры, относящиеся к виду Т. taeniatus. Можно сделать и обратный опыт. Мы можем трансплантировать небольшой участок эктодермы раннего зародыша Т. cristatus, взятый, например, с брюха, на спинную область, лежащую над крышей первичной кишки зародыша Т. taeniatus, т. е. на место нормальной нервной пластинки. Несмотря на свое происхождение от брюшной эктодермы, этот кусочек дает под влияним тканей и формообразовательных веществ хозяина, согласно своему новому положению, начало части центральной нервной системы. Однако даже и в очень малом кусочке ткани, развивающемся под всесторонним влиянием окружающей массы тканей хозяина (Т. taeniatus), проявляются все специфические свойства дифференцированной нервной ткани Т. cristatus. To же самое касается не только видовых, но и расовых признаков и мутационных изменений — их специфика выявляется всегда уже в процессах самодифференцирования при взаимодействии ядра и плазмы.
Нас, однако, интересует в большей мере вопрос о возможности изменения основ организации, создаваемой в процессах взаимодействия частей. Возможны ли и происходят ли на самом деле наследственные изменения в градиент-системах и в индукционных системах? В чем эти изменения состоят и как мутации «генов» или перестройки хромосом могут привести к принципиально новым дифференцировкам в процессе эволюции организмов? Последний вопрос разбирается подробно в третьей части книги. Здесь же мы фиксируем свое внимание на вопросе о фактическом наличии материала для такой эволюции и на его характеристике.
В настоящее время феногенетика накопила уже достаточно большой материал, показывающий, что процессы мутирования могут затрагивать все виды формообразовательного взаимодействия частей развивающегося организма, начиная с градиент-систем яйца (с его организацией плазмы), зародышевых листков и зачатков органов PI кончая индукционными системами и системами морфогенетических и эргонтических корреляций.
Мы уже заметили, что правое и левое вращение раковины брюхоногих моллюсков, изменяемое в процессе простой мутации, определяется структурой плазмы яйца. Изучение индивидуального развития мутантов дрозофилы с рудиментарными крыльями или уменьшенными глазами показало, что в этих случаях уменьшены уже их самые ранние зачатки. В мутациях, касающихся строения конечностей (полидактилия) или их оперения (мохноногость) у кур, эти изменения устанавливаются в процессе формирования самих зачатков. Иногда при этом можно заметить явные перестройки зависимостей.
В градиентах может меняться их уровень и интенсивность падения. Может изменяться и направление падения, т. е. расположение градиента (например, в право- и левовращающих яйцах моллюсков). Могут, наконец, появляться и добавочные градиенты. Примером последнего могут служить различные мутации, обнаруживаемые в удвоениях органов. Развитие полидактилии у кур покоится на частичном удвоении зачатка конечности у переднего его края. Лишние пальцы развиваются в зеркальной ориентации, что указывает на появление добавочного градиента с обратным направлением падения.
В системах морфогенетических корреляций и, в частности, в индукционных системах возможны самые различные сдвиги между компонентами. Возможны: 1) изменения места и времени контакта, 2) изменения во времени появления и в концентрации формообразовательных веществ, 3) изменения во времени созревания реагирующей ткани, 4) изменения уровня нижнего или верхнего порога нормальной реактивности ткани, 5) изменения в характере самой реакции. Чаще всего, по-видимому, встречаются сдвиги во времени наступления известных реакций (1, 2, 3). Гольдшмидт полагает, что все эти сдвиги зависят от изменения в скоростях течения некоторых химических реакций, регулируемых энзимами. Это представляется весьма вероятным. Однако гипотеза об энзиматозной природе самих генов и о том, что именно гены продуцируют активные вещества типа гормонов, представляется мне и недоказанной и маловероятной.
Формообразовательные вещества представляют, во всяком случае, продукты клеточного обмена, осуществляемого во взаимодействии ядра и плазмы. С изменением состава ядра с его хромосомами и «генами» изменяются и эти продукты как в отношении их количества, так и в отношении качества. Изменяются и концентрации веществ, и время наступления, и скорости течения как биохимических, так и морфогенетических реакций. Кроме этого, изменение генотипа (с его нормой реакций) связано с изменением порогов нормальной реактивности тканей (яркий пример: различные пороговые уровни температур развития пигмента в волосах местных рас горностаевого кролика, по Ильину) и с качественным изменением самой реакции (красные или коричневые глаза у разных линий бокоплава Gammarus chevreuxi, под влиянием одной и той же мутации, по Форду).
Все эти изменения, наступающие в мутационном процессе и используемые в процессе исторического преобразования организмов, довольно понятны. Труднее, однако, представить себе, откуда берутся новые дифференцировки как результат новых и притом локальных реакций. Каждая новая дифференцировка возникает, конечно, на базе имеющейся уже дифференцировки, в ее пределах. Однако причина новой дифференцировки, очевидно, не может лежать полностью в самой этой ткани. Если в ее возникновении известную роль может играть изменение реактивности тканей, то, очевидно, локальность наступающей реакции определяется взаимоотношениями (корреляциями) с соседними частями. При этом возможны не только действительно новые топографические соотношения, но и новые взаимодействия на основе прежних соотношений — в случае изменения свойств или интенсивности выделяемых морфогенных веществ (распространяемых, например, на большую область) или в случае такого изменения порога реактивности всей ткани, что концентрация веществ лишь локально (например, только на месте полного контакта) достигает порогового уровня (или, наоборот, реакция захватывает большую область, чем в норме).
В конце концов, все сводится либо к изменению свойств и концентраций морфогенных веществ, либо к изменению реактивности тканей с их морфогенными последствиями (в том числе и установление новых соотношений). И то и другое определяется клеточной дифференцировкой и обменом веществ, получающим свою специфику в непрерывном взаимодействии между ядром и плазмой, закономерно изменяющимся в процессе индивидуального развития. Мы и в этом случае приходим к наследственным изменениям генотипа с его нормой реакций, неуклонно лежащим в основе новых дифференцировок, каким бы путем эти последние ни осуществлялись (при данных условиях внешней среды).
В особенности возможно возникновение новых дифференцировок при мутациях, связанных с приближением интенсивности морфогенных влияний к пороговому уровню реактивности тканей. В этих случаях возможны большие различия в реакции одних и тех же тканей в зависимости от ничтожных различий в биохимическом и физиологическом состоянии отдельных клеток и их комплексов. Возможно, что этим объясняется и мозаичный характер выражения некоторых мутаций. Нужно думать, что вообще новые наследственные дифференцировки «проявляются» впервые именно на базе создавшихся физиологических различий в морфологически до того однородных клеточных комплексах.
Переходя к последствиям таких изменений, следует отметить, что в результате сдвигов во времени созревания компонентов индукционных систем и изменения уровня реактивности тканей (что друг с другом связано) особенно часто наблюдаются изменения в величине зачатков и в характере их дифференцировки. Уменьшение и увеличение зачатков бывает связано с ускорением или замедлением их развития, а также с полнотой дифференцировки. Такие изменения имеют большое значение в прогрессивном и регрессивном развитии органов.
В отношении источников новых дифференцировок следует отметить значение распространения морфогенных влияний на новые области и понижения порогового уровня реактивности тканей. В результате таких изменений происходит нередко увеличение числа сходных структур и их появление в новых областях. Так, у дрозофилы известно множество мутаций с увеличенным числом щетинок и с их распространением на области, в которых нормально их не имеется. Известны мутации удвоения конечностей, удвоения щетинок. Изменчиво число яйцевых трубок и число яиц. У кур, у голубей, а также у различных млекопитающих встречается полидактилия, т. е. увеличение числа пальцев, которое представляет результат частичного удвоения зачатка конечности. У всех позвоночных весьма обычны наследственные варианты в числе позвонков как по отдельным областям, так и в целом.
Именно эти изменения лежат в основе «полимеризации», т. е. увеличения числа гомодинамов (метамеров с их органами конечностей), гомотипов (симметричных образований) и гомономов (одноименных повторностей — пальцев, лучей, каналов почки или сложной железы, щетинок, перьев, зубов и т. п.). Лишние повтор-ности развиваются, конечно, по типу существующих. Однако при различной их локализации, различном соотношении с другими частями организма и различном отношении к внешней среде имеются все данные для изменения их строения и преобразования в иные органы. Их концентрация в определенных местах может привести и к образованию совершенно новых комплексов (например, ротовой аппарат членистоногих).
Крупнейшие преобразования, лежащие в основе типов живот ного царства, основаны на увеличении числа сходных частей и различной дифференцировке гомодинамов и гомономов. Такова уже сама многоклеточность, лежащая в основе всех дифференцировок сложных животных (и растений). На этом же принципе повторности развилась многолучевая симметрия кишечнополостных животных (а затем и иглокожих).
Принцип повторности вдоль главной оси лежит в основе организации большинства червей, всех членистоногих и, наконец, в новой форме, в основе организации позвоночных животных. Однако и в основе прогрессивного развития растений лежит тот же принцип «полимеризаций». Дифференцировка слоевища на листья и стебли привела через увеличение числа этих органов к возможности дальнейшего усложнения строения и к возникновению новых дифференцировок. Преобразование побега с его листьями в сложно дифференцированный цветок представляет яркий пример возникновения нового комплексного органа на базе полимеризации, т. е. увеличения числа существующих простых органов (листьев). Кроме этого, отметим тут же, что наступление новых дифференцировок возможно также на основе имеющейся уже общей нормы реакций, при установлении новых экологических соотношений (локального характера) с факторами внешней среды, а также, в особенности, при новых функциональных преобразованиях (см. III, 3, Г).
В. ЗНАЧЕНИЕ ВНЕШНИХ ФАКТОРОВ ОНТОГЕНЕЗА
Участие внешних факторов окружающей среды в реализации унаследованных структур и их изменений бесспорно. Однако не следует переоценивать значения этой зависимости. Не может быть и речи о том, чтобы какая-либо органическая структура была полностью, т. е. в своей специфике, детерминирована внешними факторами. Любой организм, даже наиболее простой, отличается от неорганизованной материи исторической обоснованностью новых реакций. Эти реакции всегда различны для различных, хотя бы и очень близких организмов, даже при полной идентичности внешних воздействий.
Если мы устанавливаем зависимость формообразования от внешних факторов, то мы далеко не всегда, даже, быть может, лишь в виде исключения, имеем дело с детерминирующим влиянием. Можно установить различные градации этой зависимости.
а. Зависимое формообразование. О зависимом формообразовании можно говорить, когда известный фактор среды определяет если не качество реакции, то время или место ее наступления и в особенности количественное ее выражение, так что наблюдается известная пропорциональность между последним и интенсивностью данного внешнего фактора.
Такой характер имеют лишь наиболее примитивные зависимости. Мы с ними познакомились при рассмотрении влияния внешних факторов на выражение мутаций. Зависимость количественной стороны реакций от интенсивности внешнего фактора совершенно ясна для мутаций лентовидных глаз и рудиментарных крыльев. Однако эти же примеры показывают, что качественная характеристика изменения полностью определяется генотипом (их же аллеломорфы infra-bar и pennant дают обратную температурную реакцию, а нормальные аллеломорфы не дают этой реакции вовсе или дают ее в гораздо меньшей степени). Время и место наступления реакции здесь также определяется внутренними факторами. Некоторые приспособительные модификации имеют такой же характер. Зависимость поверхности листовой пластинки и толщины ее кутикулы от условий освещения, зависимость развития корневой системы от влажности почвы или высоты стебля от температуры среды (в известных пределах) выражается не только в количественных изменениях, но и в определении времени и места реакции (в пределах локализации реагирующих органов). Зависимость развития массы (поперечного сечения) мышцы от ее тренировки имеет также до известной степени количественный характер. Вместе с тем тренировка определяет и время и конкретную локализацию реакции (в определенной мышце). Таков же характер реагирования костной ткани на изменение механической нагрузки или кровеносной системы на усиление обмена веществ.
Такой относительно простой характер зависимости не составляет общего правила и не является особенно распространенным даже у растений. Обычно зависимость приобретает более сложные формы вследствие наличия системы регуляторных механизмов, ограничивающих пределы реакции более точно выработанными нормами (некоторое ограничение этих норм наблюдается и при «вполне зависимом» формообразовании; между макс, и минимальной реакциями имеется известная возможность градации изменений).
б. Авторегуляторная зависимость формообразования. Историческая обоснованность организации, характерная для всего мира организмов, связана с известной автономностью жизненных процессов и, в частности, с известной автономностью процессов индивидуального развития. Организм никогда не подчиняется пассивно влиянию факторов внешней среды. Он им активно противодействует, следуя своим собственным законам, определяемым исторически обоснованной реакционной базой. В этом случае можно говорить о регуляторных реакциях. Как в физиологических, так и в морфогенетических реакциях регуляторные процессы приобретают в процессе эволюции все большее значение. Автономность жизненных процессов получает у вышестоящих организмов все более яркое выражение.
Приспособительные модификации отдельных частей согла суются с изменениями других частей. Вырабатываются типичные приспособительные нормы значительных частей организма или всего организма в целом.
При развитии типичных форм реагирования всего организма, в виде «адаптивных норм» или типов приспособления, регулятор-ные процессы приобретают выдающееся значение. При известной интенсивности внешнего фактора, достигающей нижнего порога реактивности тканей, зависимая от него реакция наступает сразу в полном своем выражении. Дальнейшее изменение интенсивности, вплоть до верхнего порога реактивности тканей, не изменяет результата зависимых формообразовательных процессов. Между нижним и верхним порогами приспособительной реакции лежит широкий диапазон возможных изменений интенсивности внешнего фактора, на которые организм как будто вовсе не реагирует. Это указывает на падение значения внешнего фактора. Внешний раздражитель лишь освобождает на известном уровне интенсивности определенную цепь формообразовательных процессов, протекающих в основном под влиянием внутренних факторов. Повышение интенсивности внешнего фактора не оказывает заметного влияния на течение этих процессов вследствие очевидного наличия регуляторных процессов, противодействующих этим влияниям. Внешний фактор дает при достижении порога реактивности тканей организма лишь первый толчок, приводящий в действие внутренний механизм определенного комплекса формообразовательных процессов. Он не детерминирует ни качества, ни масштаба реакции. В лучшем случае (да и то не всегда) внешний фактор определяет лишь время и иногда место ее реализации. В основном он решает, на известных уровнях интенсивности, вопрос о наступлении одной или другой из возможных, исторически выработанных морфогенетических реакций. Эту форму зависимого развития мы называем авторегуляторным развитием. Оно связано, как видно, с выработкой двух или даже нескольких типичных форм, приспособленных к известным частным условиям существования. Фенотип организма оказывается полиморфным. Этот полиморфизм может иметь сезонный или экологический характер (включая биоценотический). Можно привести много примеров авторегуляторного развития, так как обычно формы приспособительных модификаций являются по существу всегда выражением этого рода зависимостей.
В особенности много такого рода авторегуляторных зависимостей имеется у растений. У водяной гречихи (Polygonum amphibium) при известной степени влажности механизм образования воздушных листьев переключается сразу на механизм развития плавающих листьев. У стрелолиста (Sagittaria sagittifolia) при некотором ослаблении освещения реакция переключается на развитие только лентовидных листьев. У многих растений образование первичных теневых листьев при известной интенсивности освещения сразу заменяется образованием световых листьев, нередко совершенно иной формы (и строения). Комбинированное влияние яркого света и низкой температуры вызывает у земляной груши развитие альпийского габитуса и т. п.
У животных такие реакции обычно не столь глубоко захватывают организацию. Весьма распространены сезонные изменения окраски. Они имеют тот же определенный авторегуляторный характер. Куколка Araschnia (Vanessa) levana дает при выдержке в температуре ниже 0° (нормальная зимовка) начало типичной весенней рыжей форме A. levana. При содержании куколки при температуре выше 2° получается темная летняя форма A. prorsa. Подобные же сезонные формы, зависящие в своей реализации от температуры (или более сложного комплекса факторов, действующих на эндокринную систему), широко распространены у птиц и млекопитающих. Периодическое созревание половых продуктов также связано у птиц нередко с температурой среды и условиями освещения (длиной светового дня).
Иногда полиморфизм животных развивается в связи с различиями в питании. Известно, что тип развития рабочей пчелы или плодовитой самки определяется именно различиями в питании. Подобным же образом развитие мужского пола у червя Вопеlliа определяется установлением паразитического питания (на самке того же червя).
Во всех этих случаях механизм развития определенной типической организации включается автоматически в результате освобождающего действия внешнего фактора (на известном уровне его интенсивности).
На базе каких наследственных изменений развиваются такие реакции, совершенно ясно. Такие наследственные изменения встречаются у всех организмов — даже у животных с автономным и мозаичным развитием. У мухи дрозофилы зависимые морфогенетические реакции характеризуют именно различнейшие мутации. У нормального «дикого» типа их нет или они мало выражены, так как они уничтожаются в процессе эволюции автономно-мозаичного организма. Мы приводили примеры ярко выраженной зависимости мутаций уродливого брюшка (Abnormal abdomen) и безглазия (eyeless) от качества корма, а также примеры зависимости выражения мутаций рудиментарных крыльев (vestigial, pennant) и удвоения конечностей (reduplicated) от температуры. У нормальных мух эти реакции не дифференцируются (они не имеют положительного значения). Однако многочисленные изменения нормы реакций (в том числе высота температурного оптимума у той же дрозофилы) являются бесспорной базой не только для развития видовых и расовых различий, но и для возникновения сезонного или экологического полиморфизма. У растений известны и расовые и индивидуальные различия в световых и температурных реакциях (световые и теневые листья, высота стебля). Известно, что влияние температуры и длины светового дня на прорастание, вегетацию и окончательное формирование и созревание бывает различным для разных рас и сортов растений. И в этом имеются индивидуальные различия. Начало дифференцирования подобных же авторегуляторных реакций видно на различных расах горностаевого кролика, где на базе общей зависимости пигментообразования от температуры развиваются некоторые типичные реакции на температурные условия среды. Они устанавливаются у разных рас на различном температурном уровне. В известных границах температур (для московской расы в пределах от 2 до 14) окраска остается постоянной (белая с черными носом, ушами, хвостом и концами лапок), что указывает на развитие регуляторных механизмов (рис. 8).

Рис. 8. Распределение температурных порогов пигментообразования в волосах горностаевого кролика. По Н. А. Ильину, 1926 — 1927
в. Автономно-мозаичное развитие. При автономном развитии роль внешних факторов снижается еще более, чем при авторе-гуляторном формообразовании. Основное значение переходит к внутренним факторам развития. Внешние факторы теряют роль пускового механизма — все морфогенетические реакции включаются под влиянием внутренних факторов. Лишь некоторые начальные процессы находятся под контролем факторов, внешних по отношению к яйцу (эндокринные факторы и взаимодействие с тканями яичника при созревании яйца, факт проникновения сперматозоида, откладка яиц и иногда значение внешних, в частности термических, влияний как стимуляторов и освобождающих факторов; последние имеют особое значение при наличии зимнего покоя яиц; реже тот же контроль простирается и на стадии личиночного развития или метаморфоза). Известный комплекс факторов, входящих в «нормальные» условия внешней среды, определяет лишь возможность нормального развития. Из детерминирующих факторов они становятся лишь условиями, допускающими в известных, иногда довольно широких, границах, изменения интенсивности (например, температуры, влажности) без нарушения нормального развития. Нижний и верхний пороги реактивности тканей на эти факторы раздвигаются шире, чем при авторегуля-торном развитии, так что они оказываются практически всегда в пределах обычных колебаний этих факторов во внешней среде.
Автономные процессы развиваются, очевидно, на базе авторе-гуляторных процессов именно по пути указанного изменения нормы реакций (широкого расхождения обоих порогов реактивности) и дальнейшего усложнения регуляторного аппарата. И у автономно-мозаичных организмов возможны наличие отдельных авторегуляторных морфогенетических процессов и даже явления модификационного полиморфизма, как это видно на примерах сезонного полиморфизма насекомых и полиморфизма колониальных форм. Что касается основной базы для эволюции автономно-мозаичных организмов и механизма реализации у них наследственных изменений, то этот вопрос уже разбирался в начале настоящей главы — в отделе А, посвященном роли ядра и плазмы (отдел Б, посвященный другим внутренним факторам развития, хотя и относится к регуляторному типу развития, но практически захватывает и мозаичные организмы, так как полной мозаичности никогда не бывает — ранние стадии развития имеют всегда более или менее ясно выраженный рсгуляторный характер).
г. Автономно-регуляторное развитие. Дальнейшее развитие регуляторных механизмов привело, в особенности у позвоночных животных, к несколько иному и притом более совершенному развитию автономности онтогенеза.
Внешние факторы оказывают здесь, правда, некоторое формообразовательное влияние, но в несколькой иной форме. У мозаичных организмов они играют иногда известную роль как стимуляторы развития яйца или зародыша; реже они оказывают модифицирующее влияние на развитие личинки или процессы метаморфоза. У регуляторных организмов с автономным развитием влияние внешних факторов сказывается только на поздних, «функциональных» стадиях развития, и даже у взрослого животного. Вместе с тем это влияние сказывается иначе.
Конечно, и у растений и у низших животных факторы внешней среды доходят до клеток организма в существенно преобразованном виде. Специфика реакций всегда зависит от самого организма и его физиологического состояния. Внешние факторы могут вызывать лишь известное уклонение в течении физиологических процессов. При авторегуляторном развитии реакции организма усложняются вмешательством регуляторных процессов, которые могут действовать против изменения течения физиологических процессов, нейтрализуя неблагоприятные влияния внешних факторов. При автономном развитии регуляторные процессы приобретают еще большее значение. В частности, через сложную систему морфогенетических и эргонтических корреляций приспособительные изменения (адаптивные модификации) отдельных органов согласуются с целой серией коррелированных изменений, захватывающих всю организацию. При этом сама адаптивная модификация создается, за немногими исключениями (касающимися, главным образом, покровов с их роговыми образованиями и пигментацией), в результате функциональной деятельности самого органа. Косвенный характер влияния факторов среды выступает здесь с гораздо большей ясностью, чем при «зависимом» развитии. Кроме того, здесь нет исторически выработанных рамок в виде обособленных «типов» приспособления или ясно ограниченных адаптивных норм. Развивается всегда лишь одна основная форма (мономорфизм); однако, при широкой возможности адаптивных реакций в отдельных органах и при существовании сложного механизма корреляций регуляторного характера эта одна основная форма претворяется в практически бесконечное число вполне гармоничных и приспособленных вариантов. Большое значение приобретает, таким образом, «упражнение и неупражнение» органов. При этом внешняя среда оказывает свое влияние на формирование организма не столько в виде физических факторов (неорганической среды), сколько в виде биоценотических соотношений, сказывающихся, прежде всего, на поведении животных при добыче пищи и защите от врагов.
Этим дается и совершенно новая база для дальнейшей эволюции (см. раздел V).
Материальная основа наследственных изменений организмов с автономно-регуляторным развитием та же самая, что была рассмотрена раньше, когда говорилось о значении ядерных структур и плазмы и, в особенности, когда разбирался вопрос о значении внутренних факторов развития, т. е. корреляционных систем в широком смысле. К этому добавляется, однако, только что упомянутое значение индивидуального приспособления организма (через его функциональную деятельность), которое создает предпосылки для наследственной перестройки организации согласно пути, проложенному адаптивной модификацией (что, впрочем, происходит и при зависимом и особенно авторегуляторном типе развития; об этом см. во II разделе о стабилизирующей форме отбора, а также подробнее в моих книгах «Организм как целое» и «Пути и закономерности эволюционного процесса»). Возможно возникновение новых изменений на пути функциональной дифференциации органов, обладающих известной исторически обоснованной нормой общих реакций. Общие реакции дифференцируются на частные, а результаты этих частных реакций, имея в данных условиях приспособительное значение, фиксируются затем в процессе стабилизирующего отбора (И. Шмальгаузен, 1938а, 1942) соответствующих наследственных изменений.
6. ИЗМЕНЧИВОСТЬ ЛАБИЛЬНЫХ И СТАБИЛЬНЫХ ОРГАНИЗМОВ
Под лабильными организмами мы понимаем организмы, зависящие в своем индивидуальном формообразовании от влияния различных случайных изменений в факторах внешней среды, а под стабильными — формы более устойчивые, независимые в своем развитии от таких случайностей. Ясно, что изменчивость лабильных организмов будет отражать в себе эти случайные влияния — она примет довольно широкий размах, однако в основном это — модификационные изменения. С другой стороны, изменчивость стабильных организмов, свободная от этих случайных влияний, будет в большей мере отражать влияние внутренних факторов, т. е. мутационные изменения. Впрочем, при значительном развитии регуляторных механизмов влияние мутационной изменчивости может быть в значительной мере погашено.
К лабильным формам относятся в основном организмы с зависимым и авторегуляторным типами развития. Изменчивость их, как сказано, может быть весьма значительной. Так как она основывается на многочисленных влияниях случайного характера, то ее результат может быть выражен типичной вариационной кривой. Исследования Иоганнсена над изменчивостью чистых линий самоопыляющихся растений ясно показывают закономерности и широкий размах таких изменений. При типично зависимом развитии изменчивость имеет флюктуирующий характер — имеются все переходы между крайними типами (по величине, числу, интенсивности и т. п.). При авторегуляторном развитии изменчивость имеет более или менее прерывистый характер, так как она отражает скачкообразные переходы от одной адаптивной нормы к другой. Вместе с тем, вследствие наличия регуляторных механизмов, организация оказывается более устойчивой, так как в ней в меньшей мере сказывается влияние случайных изменений в факторах внешней среды.
Наследственная изменчивость выражается у лабильных форм в основном в различной реактивности организма, т. е. в различиях пороговых уровней, в интенсивности реакции и отчасти в качественных ее различиях. Накладывающиеся на нее модификацион-ные изменения в значительной мере скрывают ее и делают отдельные мутации незаметными. Поэтому иногда считают, что вариационная кривая, построенная по каким-либо признакам организмов известной популяции или группы особей, выражает лишь модификационную изменчивость. Это, конечно, неверно. Однако выделить из этих изменений наличные мутации и их комбинации можно только в экспериментальных культурах отдельных линий.
Лабильность форм не исключает известной стабильности. Именно организмы с авторегуляторным развитием, хотя и реагируют на изменения во многих факторах внешней среды, однако они не столь чувствительны в этом отношении, как организмы с вполне зависимым развитием. Они требуют для реализации известной адаптивной реакции определенной интенсивности воздействия. Более низкая его интенсивность не оказывает заметного влияния, а более высокая (превышающая нижний порог реактивности) не меняет заметно характера реакции. Случайные, кратковременные изменения в факторах внешней среды в большинстве случаев остаются без морфогенетической реакции со стороны организма (отвечающего лишь легко обратимой физиологической регуляцией). Адаптивные нормы хотя и разнообразны (полиморфизм), но в своем типичном выражении относительно устойчивы. Модификационный полиморфизм является высшей формой индивидуальной приспособляемости организмов с авторегуляторным развитием.
Эта индивидуальная приспособляемость лабильных организмов имеет исключительное значение в гетерогенной среде, т. е. в условиях закономерно встречающихся изменений в факторах среды. Это прежде всего различные локальные условия, в которые попадает организм, лишенный средств передвижения. Семена растений, разносимые ветром или животными, попадают именно в самые различные условия прорастания и развития. В особенности значительны различия в почвах, во влажности и освещении. Это и приводит к образованию экологического полиморфизма (экофены Турессона), столь распространенного именно у растений. Однако и сидячие животные — губки, кораллы, актинии и гидроиды — столь лабильны в своем оформлении, что это создает большие затруднения для систематиков, вынужденных пользоваться в качестве диагностических такими признаками, как строение спикул у губок, строение нематоцистов или гонофоров у гидроидов. Велика также модификационная изменчивость скелета у мшанок, сидячих аннелид и моллюсков (Vermetus, Ostraea, Chaта и др.).
Изменения в факторах среды могут иметь периодический — сезонный характер. Это приводит тогда и у растений и у животных к образованию сезонного полиморфизма. В данных условиях существования лабильность, т. е. индивидуальная приспособляемость, организации имеет решающее жизненное значение для организма.
Стабильные организмы, к которым относятся животные с автономным формообразованием, отличаются гораздо большей индивидуальной устойчивостью форм, и в особенности это касается животных с мозаичным развитием (нематоды, кольчатые черви, членистоногие,— особенно высшие,—моллюски). Стабильность организмов определяется в своем историческом развитии преобладающим значением случайных и кратковременных изменений Е факторах внешней среды (а не закономерных — локальных или периодических, которые определяют лабильность организации). Она не исключает индивидуальной приспособляемости форм и, следовательно, некоторой лабильности. Однако эта приспособляемость имеет главным образом физиологический характер и не приобретает форм экологического полиморфизма.
Изменчивость «мозаичных» организмов характеризуется наиболее ясным выражением мутационных уклонений, которые здесь вскрываются легче, чем при других типах индивидуального развития (хотя достоверное суждение о них возможно также лишь на основании результатов экспериментального анализа). Изменчивость имеет более или менее легко устанавливаемый дискретный характер. После гибридизации особей, относящихся к разным популяциям, она резко возрастает во втором поколении гибридов. Так как мутационная изменчивость покоится также на случайных явлениях, то суммарная вариационная кривая имеет тот же вид, что и кривая модификационных изменений. При резкой элиминации некоторых весьма обычных мутаций она может приобрести асимметричную форму. Изучение генетического состава популяций дрозофилы показывает их насыщенность разнообразными мутациями. Мутации легко выявляются в экспериментальных культурах и имеют ясно выраженный прерывистый характер. Это отражает качественные отличия мутаций, скачкообразно возникающих из «нормальной» организации «дикой» формы. Это не значит, что такие мутации не имеют места у лабильных организмов (наоборот, их насыщенность мутациями, вероятно, даже выше). Однако именно потому, что организация дикой «нормы» весьма стабильна, отдельные мутации ясно выделяются на этом устойчивом фоне в виде точно характеризуемых уклонений.
Биологическое значение стабильности состоит, очевидно, в определенности требований экологически ограниченной гомогенной среды. Подвижный организм занимает одну определенную экологический) нишу и «дикий» тип вполне приспособлен к этим условиям существования. Наличные колебания в физических факторах среды — температуре, влажности, качестве корма — имеют случайный и кратковременный характер. Если бы организм реагировал на эти изменения, это было бы неблагоприятно, так как изменения в ранних стадиях развития привели бы к уклонениям в организации взрослой формы, которые, в силу нового изменения в факторах среды (так как они случайны и кратковре-менны), уже потеряли бы свое приспособительное значение. Между тем, взрослый организм (с мозаичным развитием, и особенно у членистоногих) уже не изменяется. Реакция развивающегося организма, если бы она даже имела для данной стадии адаптивный характер, быстро утратила бы свою адаптивность и привела бы взрослую форму к гибели. Можно говорить об «ошибочности» реакции на кратковременные изменения среды. Стабильность организации, как следствие автономного развития, является как бы страховкой от таких ошибочных реакций.
Все же это не означает несовместимости стабильности организации с приспособительными реакциями (т. е. с известной лабильностью). У мозаичных организмов возможны сезонные изменения, как более сложные авторегуляторные процессы, индуцируемые обычно еще в предыдущем сезоне.
Кроме сезонного полиморфизма, бывает и экологический. Однако в этом случае он имеет характер генетического полиморфизма или гетероморфизма, т. е. он основывается на постоянном выщеплении вполне устойчивых форм, приспособленных к известным частным экологическим условиям. Особи сами находят для себя подходящую экологическую нишу. Кроме этого, возможны многочисленные индивидуальные приспособления функционального характера у организмов с автономным, но регулятор-ным развитием. Эти процессы отличаются относительно легкой обратимостью и характеризуют главным образом поздние стадии развития и даже взрослую форму.
Изменчивость регуляционных организмов характеризуется вновь более или менее скрытым мутированием. Изменчивость имеет в этом случае такой же флюктуирующий характер, как и при зависимом развитии лабильных организмов. Однако развитие автономно и организация все же весьма стабильна — она очень мало зависит непосредственно от изменений в факторах внешней среды, особенно от уклонений в физических факторах. Все эти изменения сказываются лишь косвенно через приспособительное поведение животного, соответственно данной обстановке (главным образом способы добывания пищи, защиты от врагов и в меньшей мере — защиты от неблагоприятных влияний климата), и через изменение функций его органов.
Биологическое значение стабильности регуляционного типа выражается в том, что устойчивый организм, защищенный в своем формообразовании от внешних влияний случайного характера (как и мозаичный организм), может, однако, активно менять среду обитания и приспособляться даже к совершенно новой обстановке. В этом случае нет предустановленных типичных приспособительных норм (как при авторегуляториом развитии) — организм по существу мономорфен (за исключением вполне возможных и здесь сезонных изменений). Вместе с тем одна и та же форма может быть реализована в бесконечном числе вариантов. Важнее всего, что отдельное изменение, установившееся путем функционального приспособления известного органа к новой обстановке, немедленно согласуется через посредство функциональных корреляций с целым рядом других изменений во всей организации. Результаты индивидуального приспособления имеют такой же целостный характер, как и при авторегуляториом развитии, но вместе с тем они не ограничиваются существованием лишь двух-трех исторически выработанных типичных норм, а устанавливаются для каждой особи в новой и все же вполне гармоничной форме. В стабильном организме регуляторного типа в индивидуальном развитии реализуется вполне устойчивая и целостная форма, не связанная, однако, какими-либо строго фиксированными стандартными нормами.
Стабильность организации означает существование более или менее сложного регуляторного аппарата, защищающего нормальное формообразование от возможных нарушений со стороны случайных уклонений в факторах внешней среды. Так как формообразовательные процессы едины и определяются, прежде всего, внутренними факторами развития, тот же регуляторный аппарат защищает нормальное формообразование и от некоторых случайных уклонений во внутренних факторах, т. е. от мутаций. Поэтому многие малые мутации в стабильном организме не проявляются. С другой стороны, однако, если мутация проявляется, т. е. получает заметное выражение, то это означает, что регуляторный аппарат оказался недостаточным или нарушенным этой мутацией. В обоих случаях это ведет к уменьшению стабильности, т. е. означает некоторую лабилизацию формообразования. Поэтому мутации феногенетически всегда менее стабильны, чем исходная норма (рис. 2, 3, 22, 25). Каждая проявившаяся мутация не только способствует выявлению новых мутаций, даже незначительных, но и вскрытию имевшихся уже мутаций, которые лишь не проявлялись в нормальном организме. Это имеет большое значение в эволюции при всех изменениях внешней среды, когда прежняя норма теряет свою приспособленность.
7. ИНДИВИДУАЛЬНАЯ ИЗМЕНЧИВОСТЬ КАК МАТЕРИАЛ ДЛЯ ЭВОЛЮЦИИ
Индивидуальная изменчивость как материал для исторического изменения организмов является, следовательно, весьма гетерогенной. Хотя еще Дарвин отметил, что основное значение в этом смысле имеет наследственная изменчивость (т. е. мутации), эта последняя получает различное выражение в различных условиях. Если основным фактором эволюции является естественный отбор, то в этом процессе решающим оказывается именно конкретное выражение индивидуальных особенностей при данных условиях окружающей среды (т. е. «фенотип» организма). Поэтому мы не можем игнорировать и значения «ненаследственных» изменений, т. е. модификаций. Если они приспособительны, то они определяют переживание данных особей и тем самым влияют на их дальнейшее существование и эволюцию. Если они не приспособительны, то они ведут к элиминации таких особей, а вместе с тем и к утрате способности к таким невыгодным реакциям у данного вида. Однако приспособительные модификации представляют результат исторического развития форм реагирования, и их изменения, а также возникновения новых форм реагирования, связанных с изменением наследственной базы, т. е. нормы реакций. Всякое же изменение нормы реакций означает мутацию. Таким образом, эволюция строится все же только на мутациях. Последние имеют всегда более или менее ясно выраженный скачкообразный характер. Они дискретны в своем возникновении, в своем выражении и в передаче своих признаков потомству. Они комбинируются, но не поглощаются. Это имеет огромное значение в процессе эволюции, позволяя создавать все новые, более выгодные, более устойчивые и жизненные комбинации. Однако и их выражение изменчиво, и не только в зависимости от условий среды, но и в зависимости от индивидуальных наследственных свойств организма. Выражение мутаций, а также их комбинаций может быть погашено и в гетерозиготной комбинации с аллеломорфной формой (нормой), и в более сложных комбинациях с другими мутациями, и в результате вмешательства регуляторных механизмов индивидуального развития организма. Оно может быть скрыто и в широких нормах индивидуальной приспособляемости организма к факторам внешней среды, и в широкой способности частей организма к взаимному приспособлению.
Это погашение выражения мутаций имеет не меньшее значение в процессе эволюции, чем дискретность их проявления. Если вторая дает возможность разнообразного комбинирования, то первое дает необходимые для этого предпосылки. Большинство мутаций в отдельности неблагоприятны. Погашение их выражения (если оно не слишком значительно) обеспечивает возможность их сохранения и накопления в популяции в скрытом виде. При возникновении удачных комбинаций они могут усилить свое выражение в том же процессе индивидуального приспособления, в переходе в гомозиготное состояние, в снятии регуляторных надстроек. При передаче изменений по наследству наиболее ответственна роль ядерных структур (хромосом), хотя результаты изменения последних и реализуются лишь в процессах взаимодействия с плазмой половых и соматических клеток. Если говорить об устойчивости организма в передаче его свойств по наследству, то за эту устойчивость ответственно во всяком случае не ядро, структуры которого относительно легко изменяемы, а только система клетки в целом, вместе с ее регуляторными механизмами (а также система корреляций развивающегося организма).
В разных условиях внешней среды значение тех или иных форм изменчивости весьма различно. При закономерных изменениях факторов гетерогенной среды, в зависимости от местных условий или по сезонам (различающимся по физическим, а следовательно и по биотическим факторам), большое значение приобретают адаптивные модификации. Тогда организация индивидуально изменчива — лабильна. Наследственные изменения могут быть мало заметными. Наоборот, при преобладании случайных и кратковременных изменений (колебаний) в факторах гомогенной, в общем, среды большое значение приобретает устойчивость формообразования, развитие регуляторных механизмов, устраняющих преждевременные — и в конечном счете ненужные («ошибочные») — реакции организма. В этих случаях организация устойчива — стабильна. Наследственные изменения могут легко выявляться (при мозаичном развитии). Это не исключает индивидуальной приспособляемости (лабильности), которая при регуля-торном развитии может быть довольно разносторонней. Однако она принимает иные формы — приспособления молодого и вполне сформированного организма к условиям его существования через соответствующее поведение и функционирование органов. Образуется много вариантов организации, в которых отдельные наследственные изменения в значительной мере погашаются.
Понятие «нормального» фенотипа и «нейтральных», положительных и отрицательных, уклонений от нормы. Так как вопросы о границах «нормального» фенотипа, о критериях «нормы« и уклонений от этой «нормы» имеют большое значение для понимания механизма естественного отбора, то нам придется остановиться хотя бы на условных определениях этих понятий. В вопросах, касающихся естественного отбора, решающими моментами, с точки зрения теории Дарвина, являются переживание особи и оставление ею потомства. Поэтому переживание и оставление потомства должны служить единственным критерием для оценки индивидуальных свойств особи как «нормальных» для данных исторически сложившихся условий среды или как положительных или отрицательных уклонений от этой «нормы». При этом «нормальный» тип, конечно, не будет ни фенотипически, ни генотипически однородным. Границы «нормы» будут столь же условны, как и границы, устанавливаемые систематиками по морфологическим признакам. Эти границы охватывают всегда известный диапазон вариаций. А так как они устанавливаются по пойманным в природе, следовательно пережившим и нередко оставившим потомство, то наша условная норма фактически окажется очень близкой к типичным формам систематиков, когда они характеризуют малые систематические категории. Понятие «нормального» фенотипа условно охватывает, следовательно, всех особей данной популяции, достигающих зрелого состояния в нормальных условиях среды и оставляющих «нормальное» потомство такой же численности (среди этой «нормы» могут быть отдельные ничем не отличающиеся особи, которые случайно не оставили после себя потомства или случайно оставили очень большое потомство,— это не меняет положения).
Отдельные особи «нормального» фенотипа отличаются друг от друга индивидуальными особенностями, покоящимися в значительной мере (при одинаковых условиях среды) на генотипических различиях, представляя результат комбинирования многих мутаций. Все эти различия между особями или уклонения от средней характеристики нормы можно назвать нейтральными уклонениями. Условно можно отнести к нейтральным уклонениям и некоторые отрицательные вариации, которые при скрещивании между собой хотя и дают потомство, но менее жизнеспособное и менее плодовитое. При условии изоляции они не могли бы поддержать свое существование, но при обычном скрещивании с «нормальными» особями они все же оставляют жизнеспособное потомство и поэтому принимают участие в формировании генотипа данной популяции. Такие условно нейтральные уклонения полностью не элиминируются и, следовательно, имеют известное значение в процессе эволюции.
Нейтральными, или безразличными, уклонениями мы называем соответственно все те вариации, которые выражаются в изменениях фенотипа, не выходящих за пределы «нормы» при нормальных условиях развития и при обычных условиях существования и скрещивания (т. е. при скрещивании с «нормальными» особями).
Положительными уклонениями от «нормы», а также положительными мутациями мы называем все те уклонения и мутации, которые при нормальных условиях развития и при обычных условиях существования оставляют после себя достигающее половой зрелости и плодовитое потомство, превышающее своей численностью исходные формы, т. е. распространяющееся в популяции (даже без повторного мутирования; вопроса случайной гибели мы здесь не касаемся).
Отрицательными уклонениями от «нормы», а также отрицательными мутациями мы будем называть все те уклонения от «нормы» и все мутации, которые при нормальных условиях развития и при обычных условиях существования, в частности при скрещивании с нормальными особями, не оставляют потомства или оставляют мало жизнеспособное или мало плодовитое потомство, рано или поздно исчезающее из популяции (т. е. численность потомства оказывается ниже численности исходных форм даже при скрещивании с нормальными особями). При наличии повторного мутирования они, конечно, не исчезают.
В составе «нормы» имеется всегда множество индивидуальных уклонений от некоторой средней, которые в значительной мере покоятся на различных комбинациях нейтральных, а также малых положительных и малых отрицательных мутаций, не выводящих отдельных особей за пределы нашей условной нормы. В потомстве «нормы» происходит непрерывное расщепление и перекомбинирование (а также и мутирование), при которых, вследствие естественной элиминации неудачных комбинаций и значительной случайной гибели, обычно лишь небольшая часть особей образует новое поколение «нормы».
Оценивая значение индивидуальных уклонений в эволюции, мы должны, в первую очередь, отметить существование принципиальной разницы между крайне редкими положительными уклонениями и условной нормой, с одной стороны, и многочисленными отрицательными уклонениями от нее, с другой. Первые, оставляя какое-то потомство, дозревающее (хотя бы в числе одной особи) и скрещивающееся с другими «нормальными» особями, могут принять какое-то участие в дальнейшей эволюции (если они сами по себе не вносят чего-либо положительного, то возможно их участие в создании благоприятных комбинаций). Вторые не оставляют никакого потомства или их потомство оказывается бесплодным и не принимают никакого участия в дальнейшей эволюции. «Нормальные» особи все преходящи, но все они оставляют какой-то след в популяции. След этот не прямо зависит от характера мутации, так как выражения последней меняются при изменении генотипа в процессе скрещиваний. Так как огромное большинство уклонений имеет отрицательный характер, то наибольшее значение в процессе отбора получает селекционное преимущество «нормы» перед отрицательными от нее уклонениями.