
Проблемы Эволюции |
М.: Академия, 2001. 425 с.
Содержание:
Из части II
Сложные приспособления и проблема их развития
Развитие приспособлений на основе преадаптаций
Морфофункциональные преадаптации и смена функций
Из части III
Происхождение многоклеточных организмов
«Взрывная эволюция» в начале кембрия
Из части IV
Эволюционные преобразования корреляционных систем
Каскадные корреляции и ключевые изменения
Причины макроэволюционного типостаза
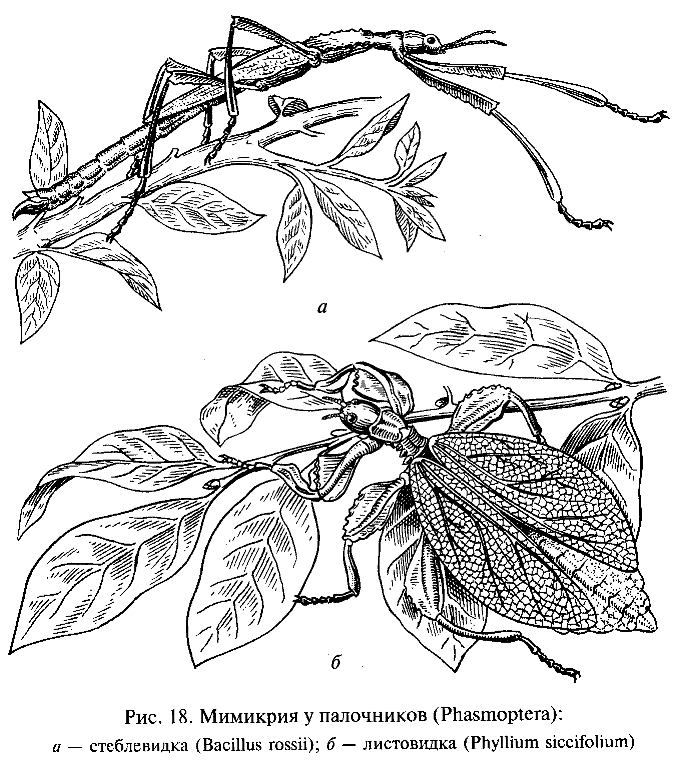
Каждый вид организмов обладает теми или иными приспособлениями, позволяющими ему существовать в определенных условиях среды. Наличие приспособлений придает строению и жизнедеятельности организмов черты целесообразности, которая всегда обращала на себя внимание естествоиспытателей. Невольно поражают воображение сложность и совершенство таких адаптации, как, например, взаимные приспособления цветковых растений и опыляющих их насекомых. Сюда относятся характерные особенности строения и окраски венчика цветов и соответствующие особенности строения челюстного аппарата и конечностей насекомых-опылителей, а также черты их поведения. Поразительны явления мимикрии, или подражательного сходства, когда один из видов имитирует особенности внешнего строения и окраски другого, «маскируясь» под несъедобный или ядовитый для хищника объект: «подражание» листу в форме и окраске крыльев или имитация стебля растений у насекомых-палочников (Phasmoptera) (рис. 18), «подражание» неядовитых видов ядовитым или хорошо защищенным (например, уже упоминавшееся выше сходство самок бабочки-парусника с несъедобными бабочками-данаидами) и т. п.
Теория Ч.Дарвина впервые дала научное решение проблемы происхождения адаптации как неизбежного результата естественного отбора. На заре мутационизма многие ученые склонялись к мысли, что новые приспособления возникают сразу готовыми — как случайный результат мутаций крупного масштаба. Подобные воззрения позднее развивал Р. Гольдшмидт, считавший, что крупные «системные мутации» резко изменяют конституцию организма, приводя к появлению «многообещающих уродов» (hopeful monsters), которые могут случайно оказаться приспособленными к новым условиям существования. Эта концепция маловероятна как с генетической, так и с экологической точки зрения. Как уже отмечалось, сколько-нибудь значительные мутации обычно ле-тальны, поскольку они приводят к нарушениям нормальной интеграции организма на том или ином этапе онтогенеза. Маловероятно возникновение посредством крупной мутации такого нового фенотипа, который оказался бы не только жизнеспособным, но и плодовитым, а к тому же априорно приспособленным к каким-либо конкретным условиям существования. В связи с этим Э.Майр справедливо заметил, что организмы, которые возникнут в результате крупных мутаций, скорее окажутся «безнадежными уродами» (hopeless monsters), чем «многообещающими», по Р.Гольдшмидту.
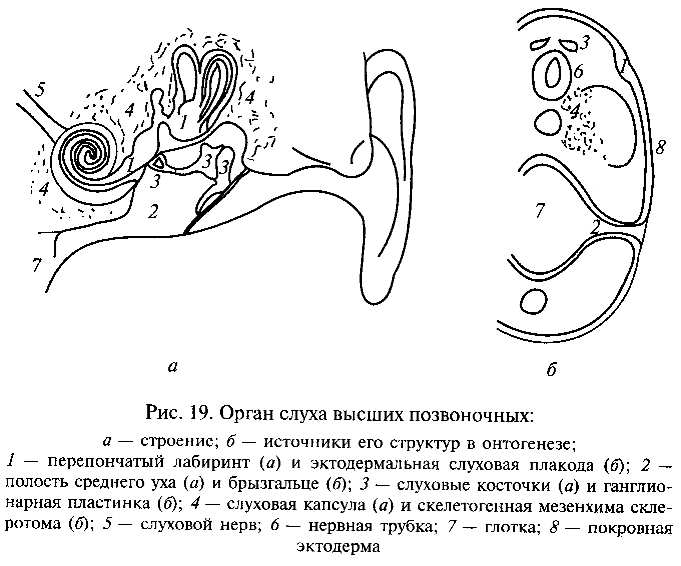
Но наиболее важным возражением против существенной роли крупных мутаций в возникновении новых типов организации является невозможность объяснить этим путем формирование сложных комплексных адаптации, образованных разнообразными компонентами, развитие которых кодируется разными генетическими системами и обеспечивается независимыми цепями морфо-генетических процессов в онтогенезе. Именно таковы многие адаптации общего значения, составляющие основной план организации высших таксонов. Например, слуховой анализатор у высших наземных позвоночных (амниот) включает структуры, имеющие разное эволюционное происхождение и формирующиеся в онтогенезе разным путем из различных зачатков (рис. 19). Так, внутреннее ухо развивается из слуховой плакоды (утолщение зародышевой эктодермы); полость среднего уха — из зачатка рудиментарной жаберной щели (брызгальца); слуховые косточки среднего уха (звукопроводящий аппарат) — из элементов подъязычной и челюстной висцеральных дуг, возникающих, в свою очередь, из материала так называемой ганглионарной пластинки эктодермаль-ного происхождения; скелет слуховой капсулы — из скелетоген-ной мезодермы головы; наконец, слуховой нерв и мозговые центры слухового анализатора формируются из невральной эктодермы. Совершенно ясно, что невозможно возникновение всей этой сложной, но целостной системы как путем случайного подбора различных мутаций (например, при дрейфе генов), так и посредством одной крупной «системной» мутации. Несомненно, естественный отбор играет решающую роль в накоплении мутаций и интеграции их фенотипических проявлений, обусловливая развитие приспособлений организмов.
Однако в этой проблеме имеется ряд сложных вопросов, и некоторые из них отчетливо представлял уже Ч.Дарвин. Первая трудность связана с тем, как отбор может контролировать начальные этапы развития нового приспособления, когда новая структура (или функция) еще столь слабо выражена, что ее небольшие изменения не могут принести организму каких-либо заметных выгод. Пытаясь разрешить эту проблему, Ч.Дарвин в некоторых случаях даже пытался привлечь «унаследование последствий употребления и неупотребления органов» (например, для объяснения развития асимметричного расположения глаз у камбал или развития цепкого хвоста у южноамериканских обезьян). Очевидно, такое объяснение было шагом назад по отношению к теории естественного отбора и не может быть признано удовлетворительным.
Особенно трудно объяснить действие отбора на первых этапах развития таких сложных органов, которые вообще могут эффективно функционировать только при достаточно высокой степени своего развития. Таковы, например, уже упоминавшиеся нами сложные адаптации — летательный аппарат птиц, звукопроводящая система среднего уха наземных позвоночных, плавательный пузырь костистых рыб, грудная клетка амниот в роли легочного воздушного насоса и т. п. Трудно понять, какую приспосо-бительную ценность для организма могут иметь первые зачатки подобных сложных органов и как на них будет действовать отбор, поскольку естественный отбор не может «предвидеть» будущего развития данного признака и его будущего значения для организма.
Путь к решению этой проблемы помогает найти концепция морфофункциональных преадаптаций. Сам термин «преадаптация» принадлежит французскому ученому Л. Кено. Кено понимал под преадаптациями такие особенности организмов, которые возникают как случайные вариации, первоначально безразличные для организма (т. е. не приносящие ему ни вреда, ни заметной пользы), но после изменения условий существования или образа жизни данного вида оказываются готовыми приспособлениями, полезными в новых условиях. Очевидно, эти представления Кено о развитии приспособлений из случайных преадаптаций без участия естественного отбора примыкают к воззрениям ранних генетиков-мутационистов. Однако в концепции Кено было все же рациональное зерно, заключавшееся в предположении, что сложные приспособления к каким-либо конкретным условиям могут первоначально развиваться в совершенно иной среде обитания и выполнять другую роль, и лишь позднее — в новых условиях — принять на себя новые, прежде им не свойственные функции. Кено справедливо указывал, что освоение новой среды обитания любым видом организмов возможно только при наличии у него таких особенностей строения, физиологии и поведения, которые дают возможность выжить в новых условиях, но возникнуть эти особенности могли только до начала освоения нового образа жизни, т. е. в прежней среде обитания. Такие особенности и являются преадаптивными.
Кено обратил внимание на то, что, в сущности, уже Дарвин подошел к идее преадаптации, анализируя в «Происхождении видов» развитие некоторых приспособлений. Так, швы между костями черепа млекопитающих облегчают прохождение головы детеныша при родах, однако наличие таких сочленений между костями черепа связано совсем не с живорождением, и возникли эти соединения костей в черепе древних позвоночных задолго до появления живорождения. «Лазающие пальмы» Малайского архипелага имеют приспособления в виде крючков, позволяющие им удерживаться на ветках и стволах деревьев, взбираясь по ним вверх. Но точно такие же крючки существуют и у многих нелазающих родственных видов растений, которым они служат как защитные приспособления.
За время своего существования концепция преадаптации претерпела значительные изменения, и ныне ее положения существенно отличаются от взглядов Л. Кено. В первую очередь это относится к отказу от наивных представлений, что преадаптации являются уже готовыми приспособлениями, возникающими во всей их сложности чисто случайно, без участия естественного отбора.
Концепция преадаптации охватывает самые различные процессы и явления, связанные с развитием у организмов новых приспособлений и принятием органами новых функций. Эти процессы и явления могут относиться к особенностям строения, физиологии, индивидуального развития, поведения и образа жизни организмов. В наиболее общем смысле преадаптациями называют такие свойства организмов, которые потенциально имеют приспосо-бительную ценность для еще не осуществленных форм взаимодействий организма и среды или для новых способов функционирования отдельных органов. Так же как и в отношении термина «адаптация», пре-адаптацией называют и состояние организма (или органа), и процесс развития преадаптивного состояния. Процесс усовершенствования уже существующих приспособлений к уже освоенной среде обитания (или — более узко — процесс совершенствования прежних способов функционирования отдельных органов) называют постадаптацией.
Современная концепция преадаптации включает два основных аспекта — генетико-экологический и морфо-функциональный. Первый из них стоит ближе ко взглядам Л. Кено.
При генетико-экологическом подходе в качестве преадаптив-ных рассматриваются такие нейтральные и «слабовредные» особенности фенотипа, которые поддерживаются мутационным процессом, эффектом плейотропии, коррелятивными взаимосвязями в организме и тому подобными механизмами и оказываются адаптивными при изменении условий внешней среды. В сущности, ге-нетико-экологическая концепция преадаптации сводится к представлениям о возможности использования видом первоначально неприспособительных признаков (возникших на основе случайных мутаций) для развития адаптации при изменениях направления отбора. Сами по себе эти представления не вызывают возражений, но, в сущности, этот подход является малопродуктивным, так как конкретный анализ развития преадаптивных признаков в этом аспекте отсутствует. Кроме того, генетико-экологическая концепция преадаптации выпускает из поля зрения творческую роль естественного отбора, за что она нередко подвергалась критике (Отметим, что взгляды сторонников генетико-экологической концепции преадаптации или даже взгляды Л. Кено и других ранних мутационистов часто критикуются в качестве «теории преадаптации», которая к этим взглядам отнюдь не сводится).
В отличие от генетико-экологической концепции, рассматривающей возникновение преадаптивного состояния на основе случайного подбора мутаций, морфофункциональная концепция, разработанная Д. Симпсоном и В. Боком, подчеркивает развитие преадаптации на базе прежних приспособлений, под непосредственным и непрекращающимся контролем естественного отбора. Усовершенствование приспособлений к существующим условиям может оказаться преадаптацией к новым условиям среды.
Морфофункциональная концепция преадаптации позволяет разрешить ту проблему, о которой мы упоминали в начале этой главы, — развитие сложных приспособлений, способных эффективно функционировать лишь будучи вполне сформированными. Такие приспособления (подобные челюстному аппарату, среднему уху, плавательному пузырю и др.) формируются в эволюции из существовавших прежде органов, имевших иное приспо-собительное значение. Соответствующие органы сначала приобретали новую функцию (дополнительную к их прежней основной функции), а затем, если новая функция оказывалась более важной, происходила смена функций органа. Принцип смены функций органов в эволюции был сформулирован А. Дорном в 1875 г. Такой путь филогенетических изменений возможен лишь в том случае, если имеется орган, подготовленный своей предшествующей эволюцией к принятию новой, прежде не свойственной ему функции, т.е. орган, находящийся в преадаптивном состоянии. Морфофункциональная концепция преадаптации анализирует эволюционный механизм смены функций органов.
В этом аспекте преадаптивное состояние организма (или отдельного органа) возникает не как непосредственный результат случайных мутаций, а на основе предшествующей приспособи-тельной эволюции. Естественный отбор постепенно совершенствует уже существующие приспособления организмов, благоприятствуя тем вариантам, которые обеспечивают более эффективное и экономичное функционирование различных органов. В некоторых случаях при этом может возникнуть как случайный, побочный эффект приспособительной эволюции преадаптивное состояние органа, когда последний получает возможность принять новую функцию в дополнение к его прежней функции. При этом орган получает и новое приспособительное значение (новую адаптивную роль). Если новая адаптивная роль достаточно важна для организма, дальнейшая эволюция данного органа будет направлена отбором по пути совершенствования этого нового приспособления (рис.20).
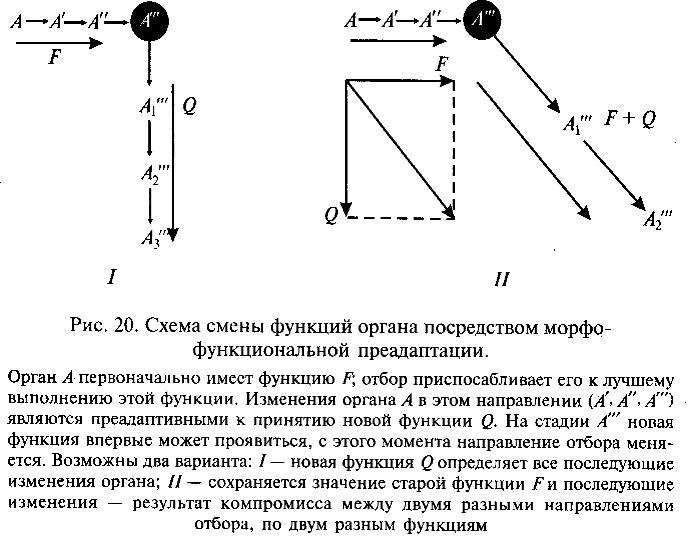
Эволюционный момент, когда орган принимает новую функцию и вследствие этого получает новую адаптивную роль, В. Бок назвал преадаптационным порогом. Прежние функция и адаптивная роль органа могут сохраниться и после принятия им новых. Тогда дальнейшие эволюционные изменения органа определяются компромиссом между двумя направлениями отбора, контролирующими старую и новую адаптации.
Важной особенностью механизма морфофункциональной преадаптации является его пороговый характер: после достижения преадаптационного порога при высокой приспособительной ценности новой адаптивной роли направление эволюционных перестроек данного органа может резко измениться. Одновременно открываются новые возможности в использовании ресурсов внешней среды, что создает предпосылки для интенсивного видообразования, приводящего к возникновению разнообразных вариантов нового конструктивного типа.
Поскольку орган подготовлен к принятию новой адаптивной роли преобразованиями, направленными на усовершенствование прежнего приспособления, т.е. постадаптациями по прежней функции, можно говорить, что преадаптация органа к новой адаптивной роли развивается на основе постадаптаций к прежней. Потребность в новом приспособлении может существовать у организмов задолго до того, как возникает подходящая преадаптация. Нужно подчеркнуть, что такая потребность далеко не всегда реализуется в эволюции. Поскольку преадаптация возникает как случайный побочный эффект приспособительных эволюционных изменений органа — очевидно, что гораздо чаще должно иметь место как раз отсутствие требуемых преадаптации, чем их развитие. Например, отсутствие предпосылок для вселения вида в новую среду обитания в целом гораздо более обычно, чем наличие требуемых для этого преадаптации.
 Целый ряд поразительных примеров смены функций, иллюстрирующих разные формы морфофункциональных преадаптации, демонстрируют эволюционные преобразования висцерального скелета позвоночных животных. У предков позвоночных (прото-краниот, по А. Н. Северцову) висцеральный скелет, вероятно, состоял из серии цельных (нерасчлененных на отделы) хрящевых колец — жаберных дуг, охватывавших передний отдел пищеварительной трубки (рис. 21) и препятствовавших спадению ее стенок. При активизации образа жизни для этих животных стала необходимой интенсификация дыхания. Последняя бьыа достигнута посредством усиления тока воды через жаберную область с развитием здесь нагнетательно-всасывающего насоса. У бесчелюстных позвоночных (предков современного класса круглоротых — Cyclostomata) жаберные дуги потеряли свою первоначальную независимость, соединившись в единую жаберную решетку продольными хрящевыми балками. При вентиляции жабер жаберная решетка, образованная гибкими хрящами, сжимается (при сокращении специальных мышц) и затем расправляется (за счет эластичности хрящевых жаберных дуг) как целое. Этот способ вентиляции жаберной области был, вероятно, связан с развитием энтодермальных жабер, расположенных внутри от жаберных дуг.
Целый ряд поразительных примеров смены функций, иллюстрирующих разные формы морфофункциональных преадаптации, демонстрируют эволюционные преобразования висцерального скелета позвоночных животных. У предков позвоночных (прото-краниот, по А. Н. Северцову) висцеральный скелет, вероятно, состоял из серии цельных (нерасчлененных на отделы) хрящевых колец — жаберных дуг, охватывавших передний отдел пищеварительной трубки (рис. 21) и препятствовавших спадению ее стенок. При активизации образа жизни для этих животных стала необходимой интенсификация дыхания. Последняя бьыа достигнута посредством усиления тока воды через жаберную область с развитием здесь нагнетательно-всасывающего насоса. У бесчелюстных позвоночных (предков современного класса круглоротых — Cyclostomata) жаберные дуги потеряли свою первоначальную независимость, соединившись в единую жаберную решетку продольными хрящевыми балками. При вентиляции жабер жаберная решетка, образованная гибкими хрящами, сжимается (при сокращении специальных мышц) и затем расправляется (за счет эластичности хрящевых жаберных дуг) как целое. Этот способ вентиляции жаберной области был, вероятно, связан с развитием энтодермальных жабер, расположенных внутри от жаберных дуг.
Совершенно иным путем происходила интенсификация обмена воды в жаберной области у предков челюстноротых позвоночных, у которых жабры имеют эктодермальное происхождение и располагаются снаружи от висцерального скелета (см. рис. 21). У этих животных сохранилась самостоятельность отдельных жаберных дуг, и каждая из них расчленилась на несколько подвижных отделов. Сокращение жаберных мышц складывает каждую жаберную дугу как гармошку,
энергично сжимая жаберную область. Этот путь эволюции оказался преадаптивным для возникновения челюстей. Передние висцеральные дуги при наличии сильных мышц, сжимающих «гармошку», получили возможность способствовать удерживанию добычи. Подчеркнем, что эта новая адаптивная роль была приобретена на основе преадаптации, связанной с усовершенствованием прежней (дыхательной) роли висцеральных дуг. Новая адаптивная роль (захват и удерживание добычи) оказалась столь важной, что прежняя роль (участие в дыхательных движениях) у передней жаберной дуги (челюстной) отступила на второй план, а затем была утрачена (рудиментарная жабра челюстной дуги сохраняется у всех рыб в виде так называемой ложножабры, теряющей непосредственную связь с челюстной дутой и находящейся у костных рыб на внутренней поверхности жаберной крышки).
Так жаберная дуга превратилась в челюсти — орган схватывания и удерживания добычи. Наличие челюстей стало одним из важнейших преимуществ, способствовавших победе челюстноротых позвоночных над бесчелюстными в борьбе за существование. Среди современных позвоночных челюстноротые составляют примерно 99,8 % от общего числа видов, а круглоротые — лишь 0,2 %. Возможно, челюсти не могли развиться у предков круглоротых из-за объединения жаберных дуг в единую жаберную решетку. В этом случае мы наблюдаем как раз такую ситуацию, когда в эволюции большой группы животных (бесчелюстные позвоночные) не была реализована биологическая потребность в развитии челюстей — важного органа добывания пищи, защиты и нападения, так как не возникло необходимой преадаптации.
После прохождения преадаптационного порога в эволюционных изменениях челюстной дуги у челюстноротых позвоночных начался период постадаптаций, усовершенствовавших приспособления к схватыванию и удерживанию добычи. Эти постадаптации привели к возникновению задней опоры челюстной дуги в области челюстного сустава на следующую сзади висцеральную дугу (подъязычную), верхний элемент которой (гиомандибулярный) стал подвеском челюстей. Однако подъязычная дуга сохранила и свою прежнюю роль — участие в вентиляции жаберной области, и у хрящевых рыб несет даже жаберные лепестки. Очевидно, здесь мы имеем пример эволюционного компромисса между двумя направлениями отбора, одно из которых требует сохранения и усовершенствования прежней дыхательной функции подъязычной дуги, другое — развития новой опорной функции. Оба эти направления отбора контролировали дальнейшие эволюционные преобразования подъязычной дуги у рыб, с одной стороны, обусловив развитие механизма жаберной крышки у костных рыб, позволившего еще более интенсифицировать вентиляцию жаберной области, с другой — вызвав появление различных механизмов подвески челюстей к черепу, связанных с разными способами добывания пищи (амфистилия, гиостилия и др.).
Рассмотренное в этом примере преадаптивное состояние органа возникло в результате усовершенствования его прежней функции; такую форму преадаптации можно назвать собственно (в узком смысле) морфофункциональной. В других случаях преадаптивное состояние может развиваться в процессе редукции органа после утраты его прежней адаптивной роли — редукционная преадаптация. Наконец, третью категорию составляют онтогенетические преадаптации, возникающие на основе эмбрионального или личиночного состояния некоторых органов, сохраняющегося во взрослом организме, в результате задержки онтогенеза на соответствующих стадиях.
Явление редукционной преадаптации можно проиллюстрировать процессом эволюционного развития звукопередающей системы среднего уха у наземных позвоночных-тетрапод. У предков земноводных возник аутостилический череп, в котором гио-мандибулярный элемент подъязычной дуги утратил прежнюю роль подвеска челюстей; кроме того, редукция жаберного дыхания сопровождалась прекращением деятельности вентиляционного механизма жаберной крышки, которая также редуцировалась. Потеряв обе свои прежние функции, гиомандибулярный элемент также подвергся некоторой редукции. Этот элемент располагается в черепе сбоку от слуховой капсулы, его отростки упираются в стенку последней и в накладные кости височной области черепа, под которыми здесь располагается полость — рудимент жаберной щели между челюстной и подъязычной дугами (брызгальце). В воздушной среде эта полость оказывается заполненной воздухом; ее наружная стенка после редукции жаберной крышки становится относительно тонкой и, вероятно, могла вибрировать в ответ на колебания окружающего воздуха, как барабанная перепонка. Гиомандибуляре, упиравшееся в эту перепонку и в стенку слуховой капсулы, передавало трансформированные колебания воздуха ко внутреннему уху. Так гиоман-дибуляре функционально превратилось в первую слуховую косточку среднего уха — слуховой столбик или стремечко. В дальнейшем постадаптации усовершенствовали звукопередающие свойства этого элемента. Вероятно, подобная же редукционная преадаптация произошла и у предков млекопитающих после возникновения вторичного челюстного сустава при последовавшей затем редукции костей первичного челюстного сустава — квадратной и сочленовной, превратившихся в наковальню и молоточек среднего уха.
Примером эволюционного использования онтогенетических преадаптации может служить сохранение эмбрионального хрящевого скелета во взрослом состоянии, характерное для некоторых групп позвоночных (хрящевые рыбы, круглоротые, отчасти бесхвостые и хвостатые земноводные). В приспособительном отношении это было, вероятно, связано с необходимостью облегчения скелета (см. ч. III) при активизации образа жизни или при выходе на сушу.
Для освоения новой среды обитания, существенно отличающейся от прежней, организмы должны обладать целым комплексом особенностей строения, физиологии и поведения, позволяющих им существовать в этой новой среде. Такое состояние называется комплексной или конституционалъной преадаптацией. Например, древнейшие земноводные, начавшие освоение суши как среды обитания, обладали таким строением конечностей, органов дыхания и кровообращения и специфическими особенностями образа жизни и поведения, вся совокупность которых была необходима для выхода на сушу из водоемов. Все эти особенности, составившие комплексную преадаптацию, развились как специфические приспособления предков земноводных — кистеперых рыб — к жизни в пресных водоемах тропического пояса, в которых частым явлением было значительное уменьшение растворенного в воде кислорода (см. ч. III). В комплексной преадаптации, в сущности, объединяются морфофункциональный и генетико-экологический аспекты преадаптации. Как указал Г.Оше, комплекс уже сложившихся морфофункциональных преадаптации для своей реализации должен быть завершен некоторыми деталями, которые могут контролироваться немногими ключевыми мутациями. Например, эти мутации могут вызвать такие изменения физиологии или поведения, которые позволят данному виду начать освоение новой среды обитания. Это и преобразует комплекс преадаптации в новое приспособление.
 В связи с этим упомянем еще один интересный пример преадаптации, замечательный тем, что он реализовался не в далеком прошлом, как упоминавшиеся выше, а в наше время. Такие особенности строения попугаев, как крупный острый клюв с изогнутым вниз надклювьем (рис. 22), крепкие острые когти, мощная мускулатура челюстного аппарата, шеи и конечностей, выглядят хорошо приспособленными для хищного способа питания. Однако все эти признаки, несомненно, сформировались как приспособления к лазанию по деревьям: при лазании клюв также используется как хватательный орган, своего рода «пятая конечность», тем более что надклювье подвижно сочленяется с черепом, и клюв как целое может активно подгибаться вниз или отгибаться вверх (что нетрудно заметить, наблюдая за лазанием крупных попугаев по решетке клетки). Подавляющее большинство попугаев — мирные птицы, питающиеся растительной пищей, главным образом различными плодами и семенами.
В связи с этим упомянем еще один интересный пример преадаптации, замечательный тем, что он реализовался не в далеком прошлом, как упоминавшиеся выше, а в наше время. Такие особенности строения попугаев, как крупный острый клюв с изогнутым вниз надклювьем (рис. 22), крепкие острые когти, мощная мускулатура челюстного аппарата, шеи и конечностей, выглядят хорошо приспособленными для хищного способа питания. Однако все эти признаки, несомненно, сформировались как приспособления к лазанию по деревьям: при лазании клюв также используется как хватательный орган, своего рода «пятая конечность», тем более что надклювье подвижно сочленяется с черепом, и клюв как целое может активно подгибаться вниз или отгибаться вверх (что нетрудно заметить, наблюдая за лазанием крупных попугаев по решетке клетки). Подавляющее большинство попугаев — мирные птицы, питающиеся растительной пищей, главным образом различными плодами и семенами.
Однако известен один вид попугаев, который действительно использовал указанные признаки при освоении хищного образа жизни. Попугай кеа (Nestor notabilis) (см. рис. 22), обитающий в горных районах южного острова Новой Зеландии, питается обычно различной растительной пищей (фрукты, нектар цветов), а также мелкими беспозвоночными животными (черви, насекомые), которых добывает из-под камней. Однако начиная с 1868 г. были отмечены случаи нападения кеа на овец, пригоняемых на горные пастбища. Хищнические наклонности проявляют отдельные особи попугаев: они расклевывают на спине овец огромные раны (до 10 см), от которых жертва быстро погибает. Мясо погибших овец поедают не только птицы-убийцы, но и другие кеа. Поскольку овцеводство появилось в Новой Зеландии после прихода туда европейцев (до этого на острове не было других млекопитающих, кроме крыс и летучих мышей), очевидно, что переход попугаев кеа к хищничеству произошел, можно сказать, на наших глазах. При этом строение клюва и когтей попугаев не претерпело никаких изменений — их использование для новой роли было пре-адаптировано прежними приспособлениями, облегчавшими попугаям лазание по деревьям и питание плодами растений. Попугаи, нападающие на овец, ничем не отличаются от других кеа. Возможно такое изменение поведения обусловлено возникновением соответствующей мутации, позволившей использовать по-новому существовавшие прежде структуры.
Необходимо подчеркнуть, что путь преадаптации — широко распространенный, но не единственно возможный способ развития новых приспособлений. Некоторые структуры, особенности физиологии и поведения могут с самого начала формироваться как «постадаптации» (здесь термин чисто условный, поскольку отсутствует период преадаптации). Условием для этого является такая приспособительная ценность уже первых зачатков новой структуры, которая достаточна для возникновения положительного давления естественного отбора, благоприятствующего новому признаку. Примером сложного органа, возникшего таким путем, является глаз, как это убедительно показал Ч.Дарвин.
торые достигли высокого совершенства в «мире малых форм». Малые размеры тела ограничивают количество клеток в организме, в частности в центральной нервной системе. Для насекомых характерна автономизация периферических отделов нервной системы (замыкание рефлекторных дуг на уровне периферических ганглиев) и преобладание наследственно закрепленных автоматизированных форм поведения. Трахейная система ограничивает также возможности экологической экспансии насекомых: при огромном разнообразии форм и адаптации насекомые совершенно не смогли освоить наиболее древнюю и обширную среду жизни — толщу морской воды. Вероятно, это связано с высокой плавучестью тела насекомых, пронизанного густой сетью воздухоносных ходов.
Вероятно, 700—900 млн лет назад на Земле появились первые многоклеточные животные и растения. У растений возникновение многоклеточного уровня организации, вероятно, произошло на основе дифференциации лентообразных колоний, образовавшихся путем бокового срастания прикрепленных нитчатых форм или благодаря делению клеток последних в двух взаимно перпендикулярных направлениях (в одной плоскости). У колоний, прикрепленных одним концом к субстрату, различные участки находились в разных условиях по отношению к падающему свету, субстрату и водной среде. В связи с этим естественный отбор благоприятствовал возникновению некоторой дифференциации частей колонии. Первым шагом было возникновение полярности колонии; на одном ее конце выделялись клетки, служившие для прикрепления к субстрату (для них характерно ослабление фотосинтеза, потеря способности к делению), на другом же конце — верхушечные клетки, интенсивно делившиеся и образовавшие своего рода «точку роста» колонии. Естественный отбор благоприятствовал приобретению клетками колонии способности делиться в разных направлениях; это приводило к ветвлению, что увеличивало поверхность колонии. Деление клеток вдоль трех взаимно перпендикулярных осей или переплетениеотдельных нитей вело к возникновению многослойного «объемного» тела. В процессе его дальнейшей дифференциации сформировались многоклеточные органы, выполнявшие разные функции (фиксация на субстрате, фотосинтез, размножение). Одновременно между разными клетками растения складывалась определенная взаимозависимость, что, собственно говоря, и знаменует достижение многоклеточного уровня организации.
У животных активный образ жизни требовал более совершенной и сложной дифференциации организма, чем у растений. Сложность организации многоклеточных животных (Metazoa) и разнообразие ее конкретных форм стимулировали разработку различных гипотез о происхождении Metazoa.
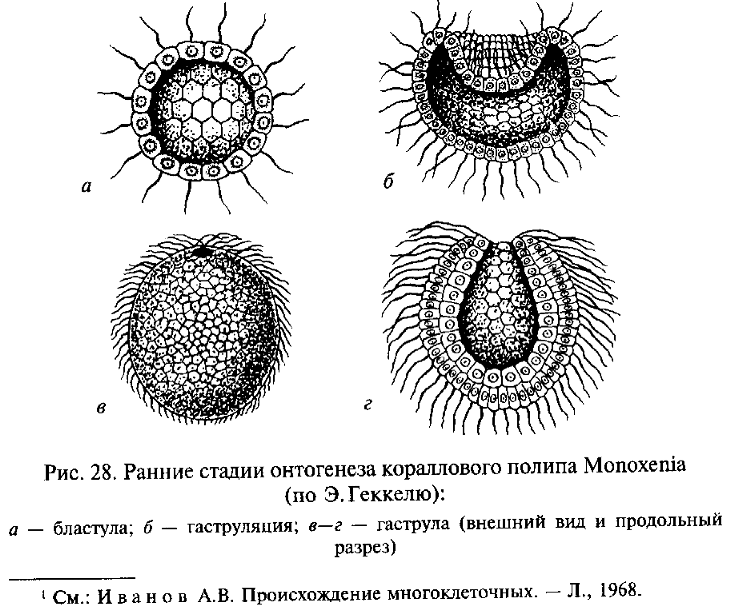
Первая из них берет начало в работах Э. Геккеля, который в разработке своей теории гастреи основывался на сформулированном им биогенетическом законе, согласно которому онтогенез данного вида организмов представляет собой сжатое и сокращенное повторение (рекапитуляцию) хода филогенеза его предков (подробнее см. в ч. IV). В соответствии с этим Э. Геккель полагал, что филогенез древнейших Metazoa в определенной степени повторяется в онтогенезе современных низших многоклеточных животных (рис. 28). Согласно Геккелю, предками Metazoa были колониальные простейшие, обладавшие сферическими колониями с однослойной стенкой, подобными бластуле — одной из ранних стадий эмбрионального развития современных многоклеточных животных. Геккель назвал эту гипотетическую предковую форму «бластеей». При направленном плавании сферическая колония — бластея — ориентировалась одним полюсом вперед, как это наблюдается и у современных колониальных простейших, например у Volvox. Согласно Геккелю, на переднем полюсе колонии возникло впячивание ее стенки внутрь, подобно тому, как это происходит при инвагинационной гаструляции в онтогенезе некоторых современных Metazoa. В результате образовался многоклеточный организм — «гастрея», стенка тела которого состоит из двух слоев, экто- и энтодермы. Энтодерма окружает внутреннюю полость — первичный кишечник, открытый наружу единственным отверстием — первичным ртом. Организация гастреи соответствует принципиальному плану строения кишечнополостных (тип Coelenterata), которых Геккель и рассматривал как наиболее примитивных многоклеточных животных.
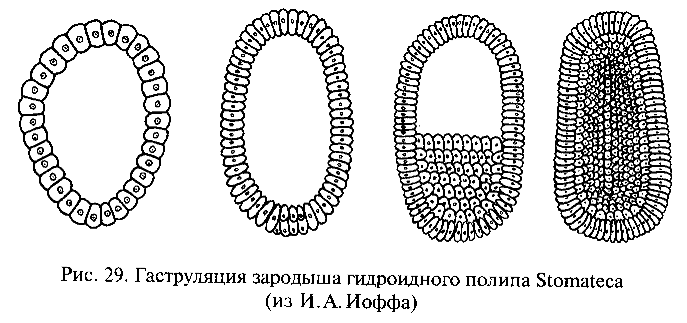
И.И.Мечников обратил внимание на то, что у примитивных кишечнополостных гаструляция происходит не путем инвагинации (впячивания одного полюса однослойного зародыша — бластулы), что характерно для более высокоорганизованных групп, а посредством миграции некоторых клеток из однослойной стенки тела внутрь (рис. 29). Там они образуют рыхлое скопление, позднее организующееся в виде стенок гастральной полости, которая прорывается наружу ротовым отверстием. Такой способ гаструляции гораздо проще, чем инвагинация, так как не требует сложного направленного и координированного смещения целого пласта клеток, и, вероятно, примитивнее инвагинации. В связи с этим Мечников модифицировал гипотезу Геккеля следующим образом. В сфероидной колонии простейших — жгутиконосцев клетки ее однослойной стенки, захватывавшие (фагоцитировавшие) пищу, мигрировали для ее переваривания внутрь, в полость колонии (подобно миграции клеток будущей энтодермы в процессе гаструляции кишечнополостных). Эти клетки образовали рыхлое внутреннее скопление — фагоцитобласт, функцией которого стало обеспечение всего организма пищей, включая ее переваривание и распределение, тогда как поверхностный слой клеток — кинобласт — осуществлял функции защиты и движения организма. Для захвата новых пищевых частиц клеткам фагоци-тобласта, по мысли Мечникова, не было необходимости возвращаться в поверхностный слой: располагаясь непосредственно под кинобластом, клетки фагоцитобласта захватывали пищевые частицы псевдоподиями, выдвигаемыми наружу в промежутках между клетками фагоцитобласта. Эта гипотетическая стадия эволюции Metazoa была названа Мечниковым фагоцителлой (или паренхимеллой); ее строение соответствует таковому паренхимулы, личинки некоторых кишечнополостных и губок. В дальнейшем как приспособление к повышению активности питания у потомков фагоцителлы произошла эпителизация фагоцитобласта с образованием первичного кишечника и возникновением ротового отверстия в том месте, где происходила преимущественная миграция клеток внутрь. По мнению некоторых ученых, это место, вероятно, соответствовало заднему по направлению движения полюсу тела, где при плавании возникают завихрения водяного потока, и поэтому условия наиболее благоприятны для захвата пищевых частиц. Гипотеза Мечникова, как и гипотеза Геккеля, рассматривает в качестве наиболее примитивных многоклеточных животных кишечнополостных и губок.
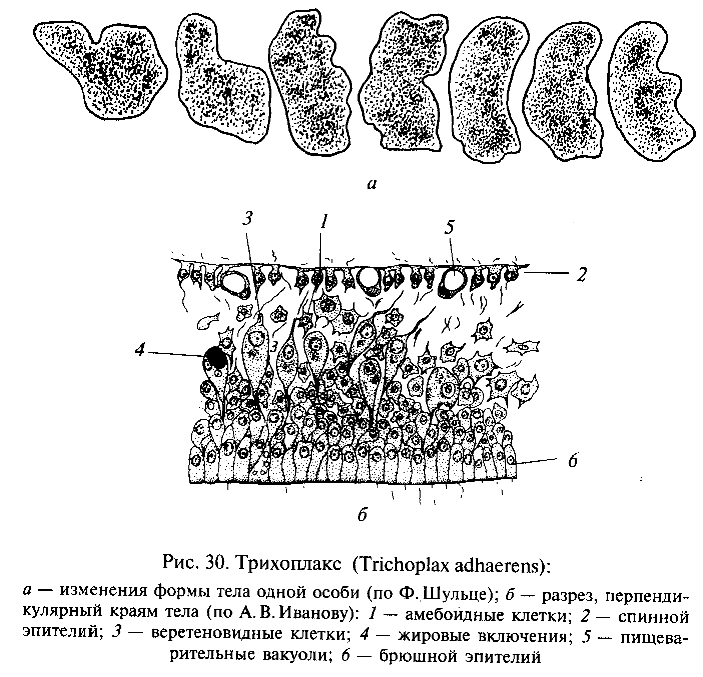
Важные сведения для понимания ранних этапов эволюции Metazoa были получены при изучении крайне примитивного многоклеточного животного трихоплакса (Trichoplax adhaerens), обнаруженного в Красном море Ф.Шульце еще в 1883 г., но детально исследованного лишь в 1970-е гг. нашего века К. Греллом и А.В.Ивановым. Трихоплакс (рис. 30) имеет уплощенное тело, лишенное полярности. Поверхность тела, обращенная вверх, покрыта плоским, а нижняя — цилиндрическим мерцательным эпителием. Внутри, между эпителиальными слоями, соответствующими кинобласту, находится полость с жидким содержимым, в котором располагаются веретеновидные и звездчатые клетки. Эти последние можно рассматривать как фагоцитобласт. Размножается трихоплакс бесполым способом — делением и почкованием. А. В. Иванов указал, что трихоплакс представляет собой как бы живую модель фагоцителлы, и предложил выделить эту форму в особый тип животных Phagocytellozoa. По-видимому, трихоплакс подкрепляет позиции гипотезы фагоцителлы И.И.Мечникова. Однако по современным представлениям непосредственными потомками фагоцителлозой среди многоклеточных были не ки-шечнополостные, а примитивные червеобразные животные, близкие по уровню организации к плоским ресничным червям — турбелляриям.
Первые ископаемые следы жизнедеятельности червеобразных многоклеточных животных известны из позднерифейских отложений. В вендское время (650—570 млн лет назад) существовали уже разнообразные животные, вероятно принадлежавшие к различным типам. Немногочисленные отпечатки мягкотелых вендских животных известны из разных районов всех континентов земного шара, кроме еще малоисследованной Антарктиды. Ряд интересных находок был сделан в позднепротерозойских отложениях на территории России — на Кольском полуострове, в Архангельской области, на реке Мая и на Оленекском поднятии в Якутии и т. д.
Наиболее известна богатая позднепротерозойская фауна, обнаруженная в Центральной Австралии в районе Эдиакары к северу от г. Аделаида. Исследовавший эту фауну М. Глесснер считает, что она включает несколько десятков видов очень разнообразных многоклеточных животных, относящихся к разным типам (рис. 31). Большинство форм принадлежат, вероятно, к кишечнополо-стньм. Это медузоподобные организмы, вероятно «парившие» в толще воды (Ediacara flindersi, Beltanella gilesi, Medusinites asteroides и др.), и прикрепленные к морскому дну полипоидные формы, одиночные или колониальные, напоминающие современных кораллов альционарий, или морские перья (Rangea longa, Arborea arborea, Pteridinium simplex и др.). Замечательно, что все они, как и другие животные эдиакарской фауны, лишены твердого скелета.
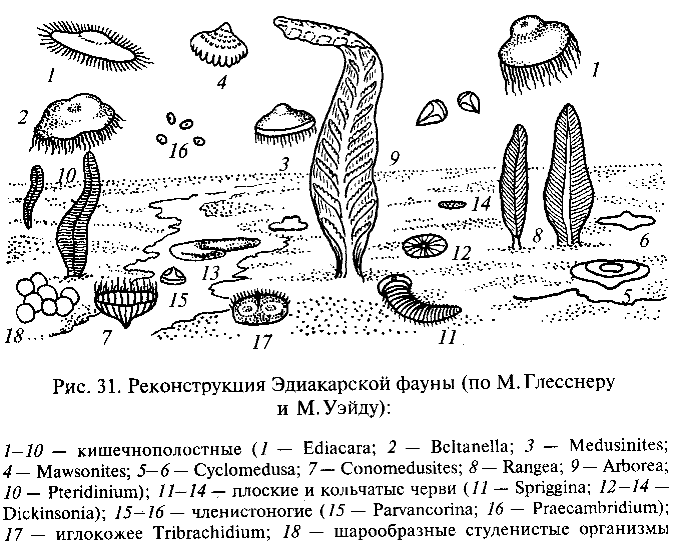
Кроме кишечнополостных в составе эдиакарской фауны найдены остатки червеобразных животных, причисляемых к плоским и кольчатым червям (Spriggina flounderi и разные виды Dickinsonia). Некоторые виды организмов интерпретируют как возможных предков членистоногих (Praecambridium sigillum, напоминающий по характеру сегментации тела трилобитов и хелицеровых) и игло-кожих (Tribrachidium heraldicum с телом дисковидной формы, на плоской поверхности которого выступают три валика, и Arkaria adami с пятилучевой звездообразной впадиной на ротовой стороне тела и с подобием амбулакральных желобков). Наконец, имеется целый ряд ископаемых организмов неизвестной таксономической принадлежности.
Многие вендские организмы были обнаружены также в вендских отложениях разных районов России: медузоподобные эдиакария и медузинитес — на полуострове Рыбачьем, птеридиниум — на севере Якутии, напоминающая сприггину вендия — в районе Яренска Архангельской области и т. д. Местонахождения вендской фауны, по богатству не уступающие эдиакарскому, были найдены на реке Сюзьма на Онежском полуострове и на Зимнем берегу Белого моря. Здесь были обнаружены ископаемые остатки свыше 30 видов бесскелетных многоклеточных животных, размеры которых варьировали от 3 мм до 30 см. Среди них вероятные представители кишечнополостных, плоских и кольчатых червей, членистоногих, иглокожих, а также ряд форм, принадлежащих к каким-то неизвестным группам. Вообще родство вендских организмов с современными группами, большинство из которых достоверно известно начиная с кембрия, остается проблематичным — различия очень велики, и некоторые исследователи считают, что известные ныне вендские организмы не связаны прямым родством с более поздними кембрийскими, а представляют слепые эволюционные ветви.
М.А.Федонкин, исследовавший Беломорскую фауну вендских животных, полагает, что некоторые из этих организмов обладают признаками нескольких разных типов животных и могут представлять исходные формы, занимающие промежуточное положение. Федонкин обратил также внимание на сходство ряда вендских организмов с личиночными стадиями некоторых современных животных, хотя вендские организмы имеют значительно более крупные размеры, чем соответствующие личинки. При всем разнообразии планов строения тела у организмов вендской фауны («вендобионтов») их объединяют некоторые общие черты организации: отсутствие скелета, конечностей, вероятно, также ды-хательньк и пищеварительных органов. Многие вендобионты вели неподвижный прикрепленный образ жизни. Некоторые исследователи полагают, что вендские организмы питались осмотически через поверхность тела или же с помощью живших в их теле фото- или хемосинтезирующих симбионтов — одноклеточных водорослей и бактерий.
Хотя среди вендских животных преобладают мягкотелые бесскелетные формы, вероятно, в те времена существовали уже и немногие виды, обладавшие раковиной. Такова, например, Cloudina, имевшая простую трубчатую раковину, состоявшую из органического вещества и кальцита. Клаудина была обнаружена в карбонатных породах, которые переслаиваются с отложениями, содержащими остатки эдиакарской фауны мягкотелых животных.
Все эти данные указывают на широкое распространение фаун мягкотелых животных в вендское время. Накопление материалов по вендским ископаемым организмам позволило некоторым исследователям поставить вопрос о расширении рамок фанерозоя, с включением в его состав «эдиакария» — периода, охватывающего промежуток времени от 670 до 550 млн лет назад.
Поскольку вендская фауна столь разнообразна и включает довольно высокоорганизованных животных, очевидно, что до ее возникновения эволюция Metazoa продолжалась уже очень долго. Вероятно, многоклеточные животные появились значительно раньше — где-то в промежутке 700—900 млн лет назад (недавно появились первые данные об ископаемых остатках многоклеточных животных, найденных в Канаде и в Китае в породах, имеющих такой геологический возраст и отделенных от пород с остатками эдиакарских организмов слоями ледниковых отложений (тиллитов)).
Таким образом, в позднем протерозое (600-650 млн лет назад) уже существовали такие группы многоклеточных животных, как губки, кишечнополостные, плоские и кольчатые черви и даже, возможно, предки членистоногих. Судя по достигнутому уровню организации, можно предполагать, что к этому времени обособились также эволюционные стволы нитчатых червей (тип Nemathelminthes), предков моллюсков и предков вторичноротых животных — олигомерных червей.
Докембрийский филогенез Metazoa можно гипотетически представить следующим образом (рис. 32). От колониальных жгутиковых (по мнению ряда авторов — от гетеротрофных форм, принадлежавших к отряду Protomonadida) путем дифференциации и интеграции колонии, с миграцией внутрь колонии клеток фагоцитобласта на заднем полюсе тела, возникли первые многоклеточные животные, организация которых соответствовала фа-гоцителле (по И. И. Мечникову). Мало изменившимися потомками этих древнейших многоклеточных являются современные Phagocytellozoa (Trichoplax adhaerens). Примитивные многоклеточные были свободноплавающими (за счет работы мерцательного эпителия — кинобласта) животными, питавшимися различными микроорганизмами — простейшими и одноклеточными водорослями.
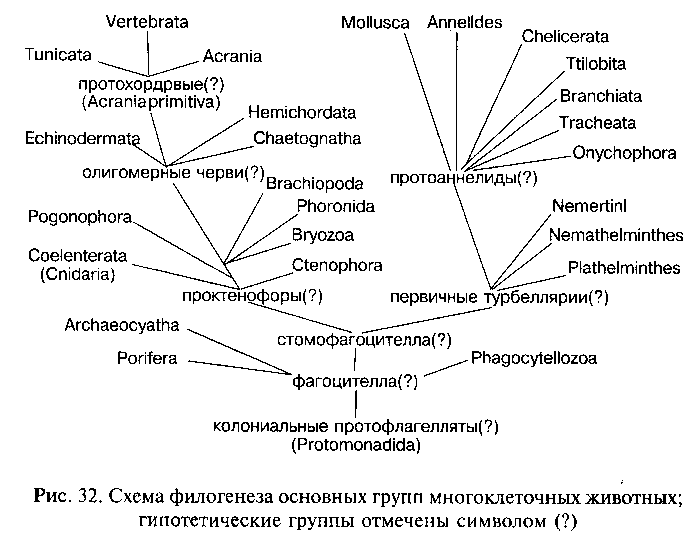
При дальнейшем развитии приспособлений к активному питанию происходила постепенная эпителизация фагоцитобласта, т. е. преобразование рыхлого скопления клеток в организованный клеточный пласт — эпителий кишечника. Эпителизация фагоцитобласта, вероятно, началась с развития на заднем по движению полюсе тела постоянного ротового отверстия. Как отметил К. В. Беклемишев, на этой стадии филогенеза организм стал питаться как целое, а не как совокупность отдельных самостоятельно фагоцитирующих клеток. Вероятно, к этому времени сформировалась и интегрирующая организм нервная система в виде эпителиального нервного сплетения. Активное плавание требовало способности ориентироваться в пространстве и координировать работу всех органов. Для осуществления этих функций на аборальном (противоположном ротовому отверстию) полюсе тела возник нейро-рецепторный комплекс, включавший нервный ганглий, осязательные щетинки и статоцист (орган равновесия). Подобный аборальный орган имеется у современных гребневиков (тип Ctenophora), а также у свободноплавающих личинок очень многих групп животных: плоских и кольчатых червей, моллюсков, членистоногих, полухордовых, иглокожих и др. Эту гипотетическую стадию филогенеза древних Metazoa можно назвать «стомофагоцителлой» (подчеркивая эпителизацию лишь ротового отдела фагоцитобласта).
Возможно, на этой стадии филогенеза произошла первая крупная дивергенция филогенетического ствола древних многоклеточных, связанная с тем, что некоторые группы этих животных перешли к освоению морского дна, другие же продолжали совершенствовать приспособления к активной жизни в толще воды.
 Современные низшие плоские черви — бескишечные турбеллярии (Acoela) в целом сохранили тот уровень организации, который, вероятно, был характерен для древнейших многоклеточных, впервые перешедших к освоению подвижного образа жизни на дне водоемов. От вендских представителей этих турбеллярий могли возникнуть филогенетические стволы, ведущие к другим группам плоских червей, к нитчатым червям и к предкам кольчатых червей (протоаннелидам). От протоаннелид обособились, с одной стороны, предки моллюсков, с другой — предки членистоногих. У всех этих групп произошла дальнейшая дифференциация фагоцитобласта. У низших червей эпителизировалась лишь его центральная часть, что привело у плоских червей к формированию разветвленного кишечника с единым отверстием — «ртом», ведущим во внешнюю среду, а у нитчатых червей — к образованию сквозного кишечника с ротовым и анальным отверстиями. У высших групп (кольчатые черви, моллюски и членистоногие) эпителизировался весь фагоцитобласт: не только его центральная часть (энтодермальный кишечник), но и периферическая (мезодерма и ее производные). Последнее привело к развитию вторичной полости тела — целома, стенки которой образованы мезодермальным целомическим эпителием. Более примитивные представители кольчатых червей, моллюсков и членистоногих обладают характерной личиночной стадией — трохофорой. В связи с этим указанные группы иногда объединяют под названием Trochozoa.
Современные низшие плоские черви — бескишечные турбеллярии (Acoela) в целом сохранили тот уровень организации, который, вероятно, был характерен для древнейших многоклеточных, впервые перешедших к освоению подвижного образа жизни на дне водоемов. От вендских представителей этих турбеллярий могли возникнуть филогенетические стволы, ведущие к другим группам плоских червей, к нитчатым червям и к предкам кольчатых червей (протоаннелидам). От протоаннелид обособились, с одной стороны, предки моллюсков, с другой — предки членистоногих. У всех этих групп произошла дальнейшая дифференциация фагоцитобласта. У низших червей эпителизировалась лишь его центральная часть, что привело у плоских червей к формированию разветвленного кишечника с единым отверстием — «ртом», ведущим во внешнюю среду, а у нитчатых червей — к образованию сквозного кишечника с ротовым и анальным отверстиями. У высших групп (кольчатые черви, моллюски и членистоногие) эпителизировался весь фагоцитобласт: не только его центральная часть (энтодермальный кишечник), но и периферическая (мезодерма и ее производные). Последнее привело к развитию вторичной полости тела — целома, стенки которой образованы мезодермальным целомическим эпителием. Более примитивные представители кольчатых червей, моллюсков и членистоногих обладают характерной личиночной стадией — трохофорой. В связи с этим указанные группы иногда объединяют под названием Trochozoa.
У тех потомков стомофагоцителлы, которые продолжали совершенствовать адаптации к жизни в толще воды, также произошла эпителизация центрального и отчасти периферического фагоцитобласта: возникла гастральная полость (первичный кишечник) и ее периферические ветви (гастроваскулярные каналы). К этому уровню организации среди современных животных ближе всего стоят гребневики, вероятно сохранившие примитивный образ жизни в толще воды. От их позднепротерозойских предков, которых можно назвать «проктенофорами», с переходом к прикрепленной жизни на морском дне возникли стрекающие кишечнополостные (тип Coelenterata, или Cnidaria).
Другие филогенетические линии, ответвившиеся от проктенофор, также осваивали морское дно, но с развитием приспособлений к активному передвижению по субстрату, подобно тур-белляриям и их потомкам, но на другом исходном уровне организации. У этих форм в результате завершения эпителизации периферического фагоцитобласта также образовалась вторичная полость тела — целом, но возникла она совершенно другим способом, чем у Trochozoa. В онтогенезе животных, происходящих от проктенофор, вторичная полость тела обособляется от первичного кишечника, как его боковые карманообразные выпячивания (первоначально было, вероятно, три пары таких выпячиваний), которые затем отшнуровываются от стенок кишки (рис. 33). Такой способ развития целома получил название энтероцельного — в отличие от схизоцельного способа, характерного для Trochozoa, у которых целом возникает в результате появления полостей внутри скоплений мезодермальных клеток, без всякой связи с первичным кишечником. О. и Р. Гертвиги и И.И.Мечников обосновали гипотезу, согласно которой энтероцельный целом возник в эволюции из гастроваскулярных каналов проктенофорных предков (энтероцельная теория происхождения целома). Энтероцельный целом характерен для типов погонофор (Pogonophora), щетинкочелюстных (Chaeto-gnatha), плеченогих (Brachiopoda), мшанок (Bryozoa) и ряда других, в том числе группы так называемых вторичноротых животных (Deuterostomia), объединяющей типы хордовых (Chordata), иглокожих (Echinodermata) и полухордовых (Hemichordata). У вторичноротых животных имеется много общего, в частности особое положение дефинитивного (присущего взрослым организмам) рта, возникающего на полюсе тела, противоположном первичному эмбриональному рту — бластопору. На месте же последнего развивается анальное отверстие. Вторичноротые, несомненно, имеют общее происхождение; в качестве их предков указывают гипотетическую группу олигомерных червей, тело которых было разделено на три отдела, имелся вторичный рот и энтероцельный целом. Среди современных вторичноротых к уровню организации олигомерных червей ближе всего стоят свободноживущие полухордовые, представителем которых является желудевый червь (Balanoglossus).
Особое положение среди многоклеточных животных занимают губки (тип Porifera, или Spongia). Эта группа характеризуется очень примитивным общим уровнем организации: губки, по существу, не имеют эпителизованного фагоцитобласта, упорядоченного внутреннего строения, настоящего кишечника, нервной системы, рецепторов и т. д. Губки отличаются от всех остальньк Metazoa чрезвычайно своеобразным онтогенезом, в ходе которого происходит инверсия зародышевых листков (экто- и энтодерма, так сказать, меняются местами). Новейшие данные молекулярных исследований показали, что губки имеют общее происхождение со всеми многоклеточными. Вероятно, они представляют собой очень раннюю боковую ветвь, обособившуюся на уровне фагопителлы. Древнейшие ископаемые остатки губок известны из вендских (эдиа-карских) отложений в Австралии.
Граница между протерозойской и палеозойской эрами отмечается резким изменением в составе и богатстве ископаемой фауны. После в основном «немых» толщ верхнего протерозоя, среди которых рассеяны относительно редкие местонахождения эдиакарской фауны, в осадочных породах кембрия (первого периода палеозойской эры), начиная с самых нижних горизонтов, внезапно возникает огромное разнообразие и обилие остатков ископаемых организмов. Среди них помимо низших многоклеточных — губок и кишечнополостных — встречаются также представители высокоразвитых типов животных — плеченогих, моллюсков, членистоногих и др. К концу кембрия появляются почти все известные типы многоклеточных животных. Этот взрыв формообразования на границе протерозоя и палеозоя — одно из самых загадочных событий в истории жизни на Земле. Благодаря этому начало кембрийского периода является столь заметной вехой, что нередко все предшествующее время в геологической истории, т. е. весь криптозой, именуют «докембрием».
Находки эдиакарской фауны и другие данные по докембрийс-ким организмам говорят о том, что эволюция многоклеточных животных началась задолго до рубежа нижнего кембрия и привела к формированию разнообразных групп. Есть основания считать, что в позднем протерозое уже сложились многие типы Metazoa. Крайняя редкость захоронений ископаемых представителей этих групп в отложениях позднего протерозоя, может быть, объясняется отсутствием у большинства докембрийских животных твердого скелета. В раннем кембрии такой скелет появился у самых разных групп животных. При этом скелетные ткани и анатомия скелета были совершенно различны у разных групп: от гибких членистых хитиновых панцирей членистоногих до монолитных известковых раковин моллюсков и плеченогих. Было высказано предположение, что причиной появления большого количества ископаемых остатков организмов в раннекембрийских отложениях было изменение гидрохимического режима водоемов, благоприятствующее скелетообразованию по чисто химическим причинам, в частности, указывают на усиленное отложение фосфоритов в морских осадках раннего кембрия. Но такое предположение не увязывается с указанным разнообразием форм и химического состава скелета у кембрийских животных. Как могло благоприятствовать повышение содержания фосфатов в воде образованию, например, кремниевой раковины, или хитина, представляющего собой комплекс азотсодержащих полисахаридов? Таким образом, едва ли правильно пытаться свести «нижнекембрийскую революцию» только к скелетизации организмов. Замечательная ископаемая фауна, включающая разнообразных животных, как наделенных скелетом, так и «мягкотелых», была обнаружена в начале 1980-х гг. в среднекембрийских сланцах Бёрджес в Британской Колумбии в Канаде. В состав этой фауны входит около 120 родов, среди которых представители губок, кишечнополостных, кольчатых червей-полихет, моллюсков, членистоногих, иглокожих, полухордовых, хордовых, плеченогих и других групп, в том числе загадочных организмов неизвестной таксономической принадлежности. Среди них выделяются крупные хищники Anomalocaris, достигавшие 2 м длины и обладавшие странным обликом: удлиненное обтекаемое тело, большие стебельчатые глаза, пара членистых околоротовых придатков, служивших, вероятно, для захвата добычи; челюстной аппарат из множества подвижных пластинок, брюшной плавник, расчлененный на ряд последовательных лопастей. В 1994 г. остатки ископаемой фауны, близкой по составу к фауне Бёрджес, были обнаружены в нижнекембрийских отложениях Южного Китая.
Для объяснения резких изменений состава и общего облика фауны и флоры в некоторые моменты истории Земли, в том числе на рубеже протерозоя и палеозоя, некоторые ученые (в частности, О. Шиндевольф) пытаются привлечь «взрыв мутаций», вызванный какими-то катастрофическими изменениями внешних условий. Подавляющее большинство мутаций снижают жизнеспособность мутантных организмов — этим пытаются объяснить массовое вымирание видов прежней фауны и флоры. В то же время общее усиление мутационного процесса, по мнению Шинде-вольфа, приводит к возникновению и быстрому распространению новых форм. При этом сразу возникают новые типы организации — скачкообразно, посредством крупных мутаций. Возможную причину такого «мутационного взрыва» О. Шиндевольф видел в повышении уровня жесткой космической радиации в результате вспышки Сверхновой звезды на достаточно близком расстоянии от Солнца.
К сожалению, эта концепция не может объяснить ни одного конкретного случая изменений фауны и флоры. Почему вымерли одни группы организмов и преуспели другие, прежде сосуществовавшие с первыми? В сущности, гипотеза «мутационного взрыва» просто подменяет анализ конкретных ситуаций постулированном универсальной причины для объяснения любого крупномасштабного эволюционного изменения. Как мы уже неоднократно подчеркивали, основой эволюционного процесса являются малые мутации, и филогенетические преобразования большого масштаба складываются из них под контролем естественного отбора. Крупные же мутации обычно детальны. Наконец, расчеты показали, что вспышки Сверхновых звезд не могут вызвать такого повышения радиации на поверхности Земли, которое имело бы предполагавшийся О.Шиндевольфом мутагенный эффект.
Вообще всегда предпочтительнее искать объяснения событиям, происходящим на Земле, не прибегая к таким гипотезам, которые постулируют универсальное влияние каких-то космических факторов, никак не опосредованное конкретными земными условиями.
В этом отношении от других концепций выгодно отличается гипотеза, выдвинутая в 60-е гг. Л.Беркнером и Л.Маршаллом, объясняющая многие важнейшие события в ходе докембрийской и послекембрийской эволюции, а также на рубеже криптозоя и фанерозоя, опираясь на закономерные изменения условий в среде обитания древних организмов. Эта гипотеза связывает развитие жизни на Земле с изменениями содержания кислорода в земной атмосфере.
В современной атмосфере Земли содержится около 21 % кислорода (это соответствует его парциальному давлению 159 мм ртутного столба). Свободный кислород атмосферы необходим подавляющему большинству современных организмов как окислитель в процессах дыхания. Водные организмы обычно дышат кислородом, растворенным в воде, но этот последний образует с атмосферным кислородом единую систему: избыток кислорода, выделяющегося в воде в результате фотосинтеза водных растений, поступает в атмосферу, а кислород атмосферы растворяется в поверхностном слое воды, по тем или другим причинам обедненной растворенным кислородом.
Однако кислородная атмосфера, столь богатая этим элементом, среди всех планет Солнечной системы присуща только Земле. Это не случайно: высокая химическая активность кислорода приводит к тому, что в условиях планет кислород в свободном состоянии долго существовать не может: участвуя в различных химических реакциях, он оказывается связанным в виде окислов и других соединений. Обилие кислорода в атмосфере современной Земли — результат фотосинтеза, осуществляемого в течение 3 млрд лет зелеными растениями. В процессах фотосинтеза из углекислого газа и воды с использованием энергии солнечного света синтезируются органические вещества (первичная биопродукция) и выделяется свободный кислород.
СО2 + Н2O + энергия солнечного света ———> С(Н2О) + О2 + 502 кДж/моль
Первичная атмосфера Земли, существовавшая во времена зарождения жизни, 3,5-4 млрд лет назад, имела восстановительный характер и состояла, вероятно, из водорода, азота, паров воды, углекислого газа, аммиака, метана, аргона и небольших количеств других газов, в том числе и кислорода. Количество свободного кислорода в первичной атмосфере не могло превышать 0,001 от современного его содержания. Это небольшое количество кислорода выделялось в результате фотодиссоциации воды ультрафиолетовыми лучами; кислород быстро входил в различные химические реакции и вновь оказывался в химически связанном состоянии.
Зарождение жизни произошло в бескислородной среде, и кислород из-за своей высокой окислительной способности первоначально был ядовит для протоорганизмов, у которых отсутствовали соответствующие защитные биохимические системы. Вероятно, протоорганизмы по способу питания являлись гетеротрофами, использовавшими в пищу различные органические соединения абиогенного происхождения, которыми, по мнению большинства ученых, бьши обогащены водоемы раннего археозоя (состояние «первичного бульона», по А.И.Опарину) и на базе которых и возникла сама жизнь. Для освобождения энергии, необходимой в жизненных процессах, первоначально использовалась анаэробная диссимиляция (брожение):
С6Н12О6———> 2СН3СН2СН + 2СО2 + 210 кДж/моль
С появлением фотосинтеза (первыми фотосинтезирующими организмами были синезеленые водоросли) в атмосферу стал выделяться кислород.
В условиях бескислородной атмосферы распространение жизни было гораздо более ограниченным, чем ныне. Дело в том, что организмы не имеют эффективных механизмов защиты от гибельной жесткой ультрафиолетовой части излучения Солнца (с длиной волны менее 250 нм). В современной атмосфере жесткая ультрафиолетовая радиация поглощается озоновым экраном — слоем озона (О3), образующегося на высоте около 50 км из кислорода (О2) под воздействием солнечного излучения и распределяющегося в основном в 15—60 км от земной поверхности. Озоновый экран надежно защищает живые организмы, которые могут существовать как в водоемах, так и на поверхности суши и в нижних слоях атмосферы. В бескислородной атмосфере раннего протерозоя озоновый экран отсутствовал, и жизнь могла развиваться только под защитой слоя воды толщиной около 10 м. Получающие наибольшие количества энергии солнечного излучения поверхностные слои воды были недоступны для организмов. Естественно, что совершенно безжизненны были и материки.
Однако фотосинтез, осуществлявшийся в океане планеты сине-зелеными, а позднее и различными группами эукариотических водорослей, в течение 2 млрд лет медленно, но неуклонно повышал содержание свободного кислорода в атмосфере. Когда содержание кислорода достигло 0,01 от современного (так называемая точка Пастера, соответствующая парциальному давлению кислорода 1,59 мм ртутного столба), у организмов впервые появилась возможность использовать для удовлетворения своих энергетических потребностей аэробную диссимиляцию. Другими словами, после достижения точки Пастера стало возможно дыхание, которое почти в 14 раз энергетически эффективнее, чем брожение:
С6Н12О6 + 6О2 ——> 6СО2 + 6Н2O + 2,87 мДж/моль — по сравнению с 210 кДж/моль, освобождающимися при брожении. Это был важнейший переломный момент в развитии жизни.
Среди современных организмов так называемые факультативные аэробы, каковыми являются многие бактерии и некоторые дрожжевые грибы, при уменьшении содержания кислорода ниже точки Пастера используют брожение, при повышении его содержания выше этой точки — дыхание (эффект Пастера).
Переход к аэробной диссимиляции в эволюции древних организмов произошел, разумеется, не сразу — для этого необходимо развитие соответствующих ферментативных систем, — но виды, приобретшие способность к дыханию, получили огромный энергетический выигрыш и в результате возможность резко интенсифицировать метаболизм и все жизненные процессы (как выяснилось, ферментная система клеточного дыхания возникла путем небольшой модификации ферментной системы фотосинтеза - см. обзор по происхождению эукариот; и впрямь, ведь эти два процесса основаны на почти одной и той же последовательности хим. реакций, только идущей в противоположные стороны; при этом надо учитывать, что практически все биохимические реакции обратимы. Это замечательный пример "смены функций" на молекулярном уровне, показывающий к тому же, что даже крупнейшие эволюционные новообразования могут возникать путем небольших модификаций чего-то уже имеющегося в организме - А.М.). Это явилось предпосылкой к дальнейшей прогрессивной эволюции и, вероятно, способствовало ускорению эволюционных преобразований.
Но достижение пастеровской точки в развитии атмосферы Земли ознаменовалось не только появлением возможности аэробной диссимиляции. При содержании кислорода в атмосфере в количестве 0,01 от современного формируется озоновый экран, который может защитить от жесткой ультрафиолетовой радиации уже и верхние слои воды в водоемах (требуется «помощь» лишь примерно 1 м воды). Это, во-первых, позволяет организмам освоить верхние слои водоемов, наиболее богатые солнечной энергией; в результате резко усиливается эффективность фотосинтеза, увеличивается биопродукция и выделение свободного кислорода. Во-вторых, чрезвычайно расширяется арена жизни: условия в водоемах значительно разнообразнее на малых глубинах, чем на больших. Освоение этого разнообразия условий в богатой энергией среде обитания неминуемо должно было привести к резкому повышению разнообразия форм жизни, к подлинному взрыву формообразования.
По расчетам Беркнера и Маршалла, точка Пастера в эволюции атмосферы Земли была пройдена примерно 620 млн лет назад; по мнению некоторых других ученых, возможно, значительно раньше—в промежутке 700-1000 млн лет назад. Но, во всяком случае, точка Пастера была пройдена в позднем протерозое, незадолго (в геологическом смысле слова) до рубежа нижнего кембрия. В этом Беркнер и Маршалл видят ключ к решению загадки ниж-некембрийского взрыва формообразования в эволюции организмов, который последовал за достижением точки Пастера в атмосфере и логически вытекает из последствий этого события (интенсификация метаболизма, освоение множества новых разнообразных местообитаний, усиление фотосинтеза, возрастание биопродукции, убыстрение эволюции).
После достижения содержания кислорода в атмосфере, равного 0,1 от современного, озоновый экран уже в состоянии полностью защитить организмы от действия жесткой ультрафиолетовой радиации. С этого момента организмы могут начать освоение суши как среды обитания. По расчетам Беркнера и Маршалла, это должно было произойти в конце ордовика (около 420 млн лет назад) (По мнению ряда других ученых, содержание кислорода в атмосфере, соответствующее 10% от современного, было достигнуто уже к началу кембрия (примерно 580 млн лет назад)). Действительно, примерно к этому времени относится появление первых наземных организмов (см. гл. 2). Современное содержание кислорода в атмосфере было достигнуто в конце пермского периода.
Гипотеза Беркнера и Маршалла привлекательна не только своей логичностью и последовательностью, но и перспективностью дальнейшего развития этих идей. Как мы уже упоминали, на рубеже нижнего кембрия у самых различных групп организмов развивается твердый скелет, облегчающий их фоссилизацию. Формирование скелета может быть также непосредственным следствием повышения содержания кислорода в атмосфере. Как показали Р. и Е. Раффы, при низком содержании кислорода в окружающей среде размеры тела многоклеточных животных не могли быть большими (вследствие низкого уровня метаболизма и энергетики организмов); газообмен с внешней средой, вероятно, осуществлялся диффузно, через поверхность тела; при этом толщина стенок тела не могла превышать нескольких миллиметров. Для таких организмов не возникало необходимости в опорном внутреннем скелете, а защитные наружные скелетные образования препятствовали бы газообмену. К. Тоув пришел к выводу, что у докембрийских животных в условиях малого содержания кислорода в окружающей среде не могло быть также хорошо развитых соединительноткан-ных образований, формирующих основу для развития скелета. Прочность соединительнотканных структур основывается на содержании в них белка коллагена, в состав которого входит аминокислота оксипролин. Ее образование возможно только при достаточно высоком содержании кислорода в окружающей среде. Следовательно, при низком содержании кислорода синтез коллагена был биохимически затруднен, и организмы не могли иметь прочных соединительнотканных структур, а поэтому и скелета, и сильно развитой мышечной системы (работа которой эффективна лишь при наличии соответствующих опорных образований).
Д. Роудс и Дж. Морзе исследовали распространение различных животных в современных водоемах с пониженным содержанием кислорода в воде (в Калифорнийском заливе и в Черном море). Была обнаружена отчетливая корреляция между содержанием кислорода в воде и характером донной фауны (бентоса). При содержании кислорода менее 0,1 мл на 1 л воды многоклеточные животные в составе бентоса отсутствуют; при 0,3—1 мл/л встречаются небольшие мягкотелые (бесскелетные) животные, зарывающиеся в ил; наконец, в более поверхностных слоях с содержанием кислорода более 1 мл/л обитают самые разнообразные животные, обладающие известковым скелетом. Эти данные представляют своего рода живую иллюстрацию к концепции Беркнера и Маршалла.
Подведем некоторые итоги. Обособление большинства типов животных произошло, вероятно, в позднем протерозое, 550—800 млн лет назад. Примитивные представители всех групп многоклеточных были небольшими лишенными скелета животными. Продолжавшееся накопление кислорода в атмосфере и увеличение мощности озонового экрана к концу протерозоя позволили животным увеличить размеры тела. Организмы получили возможность широко расселиться на малых глубинах различных водоемов, что способствовало значительному повышению разнообразия форм животных (вендская фауна).
Однако в позднем венде эта древняя фауна многоклеточных животных подверглась значительному вымиранию, которое сопровождалось новым существенным уменьшением размеров тела организмов, переживших вымирание. Возможно, этому способствовало чрезвычайно мощное оледенение (по некоторым данным, среднегодовая температура Земли понижалась в это время примерно до 5 °С). К концу венда ледники отступили, и условия вновь стали благоприятными для широкого расселения организмов.
Очевидно, виды, происходящие от общего предка и сохраняющие гомологичные гены, морфогенетические системы и структуры фенотипа, должны обладать по этим причинам значительным сходством направляющих эволюцию организменных факторов. Закономерным результатом этого является широкое распространение явлений параллельной эволюции родственных филетичес-ких линий, известное как одно из проявлений направленности филогенеза. При этом, как мы уже отмечали, гомология генов имеет существенное значение только для близкородственных видов, тогда как параллельная эволюция таксонов более высокого ранга определяется в первую очередь сходством морфогенетических и морфофизиологических факторов.
Длительной параллельной эволюции независимых филогенетических стволов в сильной степени способствуют унаследованные от общих предков каскадные взаимодействия между системами органов. Изменения ключевой системы сказываются на состоянии всего каскада зависимых от нее систем, создавая предпосылки для определенных направлений эволюционных преобразований и запрещая другие. После формирования какого-либо ключевого приспособления начинается адаптивная радиация. Возникшие при этом родственные филетические линии наследуют вместе с ключевым признаком весь комплекс обусловленных им системных взаимосвязей и соответственно весь набор эволюционных потенций, т. е. возможных направлений эволюционных изменений и эволюционных запретов. Неизбежным следствием этого является возникновение параллельных эволюционных преобразований в зависимых от ключевого признака системах организма у родственных филетических линий.
Таким образом, исторически сложившаяся организация данного таксона ограничивает для составляющих его видов выбор возможных направлений эволюции определенными рамками. Если естественный отбор длительное время благоприятствует развитию какой-либо категории адаптации (например, переработке пищи челюстями у хищников, питающихся крупной добычей, как у предков млекопитающих), направленность эволюционных преобразований в пучке родственных филетических линий получает, так сказать, «двойное обеспечение» — со стороны как организменных, так и внеорганизменных факторов. Это является причиной возникновения устойчивых эволюционных тенденций и параллельной эволюции независимых линий в филогенезе многих групп организмов.
Одним из важнейших проявлений направленности макроэволюции является поступательный характер и широкое распространение эволюционных преобразований по пути морфофизиологического прогресса. Эволюционный прогресс и связанные с ним проблемы мы рассмотрим в следующей главе.
Эволюционный прогресс является одной из центральных проблем теории эволюции, которая имеет принципиальное общебиологическое и мировоззренческое значение, будучи тесно связана с пониманием направленности эволюции жизни и места человека в природе. В то же время проблема эволюционного прогресса является и одной из наиболее запутанных. Даже в само понятие «эволюционный прогресс» разные ученые нередко вкладывают различное содержание, используя его для обозначения либо общего усложнения и усовершенствования организации, либо достижения данной группой организмов биологического процветания, либо определенной последовательности эволюционных преобразований в данном направлении (например, при развитии какого-либо приспособления). В связи с этим некоторые исследователи полагают даже, что концепция прогресса субъективна и антропоцентрична, т. е. искусственно ставит человека на вершину эволюции организмов.
Однако непредвзятый обзор общей картины развития жизни на Земле убедительно доказывает реальность происходившего в истории множества групп организмов постепенного усложнения и усовершенствования организации. Эволюция «от простого к сложному» и обозначается большинством ученых как эволюционный прогресс.
Основы современных представлений об эволюционном прогрессе были заложены А. Н. Северцовым, указавшим прежде всего на необходимость разграничивать понятия биологического и морфофизиологического прогресса. Под биологическим прогрессом понимается успех данной группы организмов в борьбе за существование, каким бы путем этот успех ни был достигнут. Биологический прогресс проявляется в увеличении численности особей данного таксона, расширении области его географического распространения и распадении на таксоны более низкого ранга (адаптивной радиации). Соответственно биологический регресс, характеризующийся противоположными показателями, означает неудачу данной группы организмов в борьбе за существование.
С другой стороны, морфофизиологический прогресс представляет собой эволюционное усложнение и усовершенствование организации. С позиций системного подхода морфофизиологический прогресс характеризуют как процесс качественной трансформации биологических систем в направлении от менее организованных форм к более организованным.
Согласно А.Н.Северцову, морфофизиологический прогресс является одним из возможных путей (наряду с другими выделенными им главными направлениями эволюции) для достижения биологического прогресса. Эволюционный «выбор» того или другого из этих путей определяется соотношением особенностей организации данной группы, ее образа жизни и характера изменений условий внешней среды.
Однако в целом ряде отношений морфофизиологический прогресс занимает особое положение среди главных направлений эволюционного процесса. Это определяется, во-первых, удивительно широким распространением арогенеза в эволюции организмов: усложнение и усовершенствование организации прослеживаются в той или иной форме в самых различных ветвях подавляющего большинства основных филогенетических стволов земных организмов. Противоположное направление — ката-генез — встречается в целом несравненно реже, чем арогенез, и катагенез всегда связан со специфическими изменениями образа жизни — переход от активной подвижности к прикрепленной или эндопаразитической жизни. Во-вторых, основные проявления морфофизиологического прогресса характеризуются у разных групп известной общностью, которую, правда, не всегда легко обнаружить среди разнообразнейших частных проявлений. Наконец, в-третьих, особой чертой арогенеза является его поступательный характер — интеграция последовательных достижений на пути морфофизиологического прогресса, как бы далеко они ни были разделены по времени (поскольку арогенез в эволюции данного филогенетического ствола обычно чередуется с периодами алло- и эпектогенеза).
А.Н.Северцов связывал морфофизиологический прогресс с повышением дифференциации организма и интенсификацией функций (в частности, с повышением интенсивности обменных процессов и энергии жизнедеятельности организма). Позднее различными учеными (И. И. Шмальгаузен, Б.Ренш, Д.Хаксли, К.М.Завадский, А. П. Расницын) к этим двум критериям арогенеза были добавлены другие. Наиболее важными из них являются: усовершенствование интеграции организма; рационализация его устройства, т. е. упорядочение организации, ведущее к оптимизации функционирования; повышение уровня гомеостаза, т. е. способности поддерживать постоянство внутренней среды организма; возрастание объема информации, извлекаемой организмом из внешней среды, и совершенствование ее обработки и использования. Другие критерии арогенеза, предлагавшиеся разными учеными, либо являются частными производными от перечисленных основных его критериев, либо не всегда коррелируют с морфофизиологическим прогрессом.
Конкретные проявления морфофизиологического прогресса в разных филогенетических стволах существенно различаются в соответствии с исторически сложившимися особенностями организации разных групп и различным характером взаимоотношений с внешней средой. Поэтому часто оказывается трудно сравнивать уровни морфофизиологического прогресса, достигнутые разными группами. Ч.Дарвин полагал даже, что «попытки сравнивать в отношении высоты организации представителей различных типов совершенно безнадежны; кто решит, кто выше, каракатица или пчела». Действительно, необходимые для подобных сравнений методы количественной оценки высоты организации еще не разработаны. Однако на качественном уровне сравнение по основным критериям арогенеза все же возможно: едва ли может возникнуть сомнение, что общий уровень организации насекомых выше, чем, например, у многоножек, и у любых членистоногих он выше, чем у кольчатых червей, а у последних — выше, чем у плоских червей, и т. п.
При этом важно подчеркнуть, что лишь совокупность названных основных критериев, не менее четырех из них, характеризует общий морфофизиологический прогресс (арогенез), т. е. повышение организации в филогенезе данной группы организмов. Арогенез происходил, например, в филогенезе позвоночных, членистоногих, моллюсков; среди таксонов меньшего ранга общий арогенез характерен для млекопитающих, птиц, насекомых, головоногих моллюсков.
В то же время во многих филогенетических стволах происходили прогрессивные изменения лишь по отдельным основным критериям. Так, например, челюстной аппарат змей значительно сложнее дифференцирован и совершеннее интегрирован, чем у их предков — ящериц. У змей он включает значительно больше подвижных костных элементов и обслуживающих эти движения мускулов, чем у ящериц. Это связано с приспособлением змей к про-глатыванию целиком крупной добычи — в процессе глотания левая и правая половины верхней и нижней челюсти могут двигаться независимо, как бы «переступая» по телу жертвы и постепенно все дальше охватывая проглатываемый объект, причем кости змеиных челюстей несколько расходятся друг от друга, удерживаясь прочными и эластичными связками. Однако это несомненное усложнение и усовершенствование челюстного аппарата змей по сравнению с ящерицами все же нельзя расценивать как проявление общего морфофизиологического прогресса, так как по своему значению для организма в целом эти эволюционные достижения имеют частный характер: общий уровень организации змей не выше, чем у ящериц. Об этом свидетельствует и уровень их энергетических и обменных процессов, и сходство гомеостаза, рецепторных систем, высшей нервной деятельности и др.
Даже столь важное проявление общей интенсификации функций организма, как повышение уровня обменных и энергетических процессов, может происходить в отрыве от других аспектов морфофизиологического прогресса и в этом случае само по себе не приводит к общему повышению уровня организации. Так, среди ракообразных наиболее интенсивным обменом веществ обладают щитни (Triops cancrifbrmis), сохраняющие примитивную и очень консервативную организацию — эти рачки являются одним из примеров персистирующих форм, поскольку их строение не претерпело заметных изменений с триасового периода (т. е. около 200 млн лет).
Для ситуаций, когда совершенствование организации происходит лишь по отдельным критериям, целесообразно использовать термины частный прогресс (предложенный А. Л. Тахтаджяном) и адаптационный прогресс (Н.В.Тимофеев-Ресовский и др.). В зависимости от степени воздействия возникающих изменений на организм в целом и от широты формирующихся адаптации, частный прогресс соответствует либо эпектогенезу (развитие эпекто-морфозов, подобных аппарату передвижения и кинетическому черепу змей), либо аллогенезу (развитие алломорфозов: ядовитые желез у змей, различных типов зубной системы у разных групп млекопитающих и т. п.).
Как уже отмечалось, А.Н.Северцов назвал эволюционные преобразования организации, ведущие к морфофизиологическому прогрессу, ароморфозами. Согласно А. Н. Северцову, ароморфозы — это такие изменения строения и функций органов, которые имеют общее значение для организма в целом и поднимают энергию его жизнедеятельности на новый качественный уровень. Как следует из приведенных самим А. Н. Северцовым примеров, конкретное содержание ароморфозов не сводится лишь к энергетическому аспекту совершенствования организации, но охватывает любые морфофизиологические преобразования, соответствующие указанным основным критериям арогенеза (дифференциации, интеграции, рационализации и оптимизации, интенсификации функций, повышению уровня гомеостаза, возрастанию усваиваемой информации и усовершенствованию ее обработки в организме). Так, несомненными ароморфозами в эволюции позвоночных животных были: развитие механизма активной вентиляции жабер (жаберного насоса) у древнейших позвоночных посредством движений висцерального жаберного скелета, приобретение челюстного аппарата (с перестройкой передних жаберных дуг), интенсификация вентиляции жабер при развитии жаберной крышки у костных рыб, приобретение последними плавательного пузыря — гидростатического аппарата, позволяющего рыбам регулировать свою плавучесть; развитие у предков высших наземных позвоночных — амниот зародышевых оболочек (амниона, серо-зы, аллантоиса), обеспечивающих возможность откладки яиц на суше; развитие мощного всасывающего (разрежающего) дыхательного насоса грудной клетки у рептилий; формирование летательного аппарата у птиц; развитие живорождения и выкармливания детенышей молоком у млекопитающих; совершенствование головного мозга у птиц, млекопитающих и человека.
Ароморфозы имеют очень высокое общее приспособительное значение, повышая независимость организма от внешней среды. В связи с этим ароморфозы ставят организмы в качественно новые отношения с внешней средой, позволяя осваивать ее ресурсы на новом уровне. Поэтому ароморфозы сохраняются в филогенезе длительное время, становясь признаками крупных таксонов. Возникающие при этом частные приспособления различных видов не затрагивают общих ароморфных признаков.
Поскольку развитие ароморфозов открывает новые возможности для освоения ресурсов внешней среды, некоторые ученые пытались, так сказать, экологизировать концепцию арогенеза, т. е. придать ароморфозам экологическое истолкование или найти экологические критерии арогенеза. В современной научной и учебной литературе можно встретить определения ароморфозов как таких изменений организации, которые позволяют освоить новую адаптивную зону (т. е. новую среду обитания или новый способ использования ее ресурсов). Иногда при этом подчеркивают, что новая адаптивная зона должна быть шире, чем у предковой группы. Однако при таком подходе, в сущности, искажается сущность концепции А. Н. Северцова, согласно которой ароморфозы — это прежде всего морфофизиологические усовершенствования, повышающие общий уровень организации, т. е. ведущие к морфофизиологическому прогрессу. В предыдущей главе было показано, что освоение новых адаптивных зон, в том числе и более широких, чем исходные, отнюдь не обязательно связано с ароморфозом и может быть достигнуто также на пути эпектогенеза или даже катагенеза.
Между разными типами изменений организмов, связанных с различными направлениями эволюционного процесса, нет резких границ. Более того, преобразования общего значения (т. е. аро-и эпектоморфозы) обычно развиваются в филогенезе из частных приспособительных изменений — алломорфозов. Так, одним из важнейших ароморфозов в эволюции позвоночных бьыо развитие легких — органа воздушного дыхания, позволившего освоить сушу как среду обитания и (у высших наземных позвоночных) резко повысить уровень метаболизма, вплоть до достижения гомойотер-мии. Напомним, однако, что легкие возникли у древних хоановых рыб как дополнительный орган дыхания, который позволяет компенсировать дефицит растворенного в воде кислорода, периодически возникавший в специфических местообитаниях этих рыб. В этих условиях легкие сформировались как типичный алломорфоз. У потомков древних кистеперых рыб — примитивных амфибий, начавших осваивать сушу как среду обитания, легкие стали основным органом дыхания. Для земноводных усложнение строения легких и преобразования подъязычного аппарата в роли дыхательного насоса приобрели значение эпектоморфоза. Эти преобразования оказали глубокое влияние на всю организацию земноводных и открьыи для них доступ в новую обширную адаптивную зону, но не привели к существенным изменениям общего уровня организации, который у земноводных по всем основным критериям в целом не выше, чем у высших костных рыб. Наконец, для высших наземных позвоночных — амниот — высокая степень дифференциации легких и дыхательных путей и возникновение механизма всасывающего дыхательного насоса грудной клетки стали важным ароморфозом, способствовавшим значительной интенсификации газообмена и общему повышению уровня организации. Таким образом, как показал Б.С.Матвеев, один и тот же орган может иметь значение и частной адаптации, и адаптации общего значения — в зависимости от степени своего развития и общей роли в организме. В эволюции алломорфоз может преобразоваться в эпектоморфоз, а этот последний — в ароморфоз.
Особое положение морфофизиологического прогресса среди разных направлений макроэволюции определяется прежде всего его поступательным характером, т. е. интеграцией последовательных ароморфозов, ведущей к постепенному общему повышению уровня организации. Эта особенность и придает эволюции по пути арогенеза черты направленности. Вопрос о причинах поступательного характера арогенеза остается одной из центральных проблем теории макроэволюции.
Ламарк, которому принадлежит заслуга первооткрывателя прогрессивной эволюции («градации»), рассматривал ее как проявление имманентных свойств организмов, изначально заложенных в них Творцом. Идеалистическое понимание эволюционного прогресса разделяли такие крупные ученые, как К. фон Бэр, Г. Ф. Осборн, Э.Коп, Л.С.Берг, П.Тейяр де Шарден и др. Основой всех идеалистических концепций эволюционного прогресса является допущение изначально присущих организмам стремления и способности развиваться в направлении усложнения и совершенствования, далее не анализируемых и представляющих собой своего рода имманентные метафизические сущности.
Примером подобных идей может служить концепция П. Тейяра де Шардена, согласно которой любая энергия имеет психическую природу. В каждом материальном элементе энергия делится на две составляющие: «тангенциальную», обеспечивающую взаимодействия друг с другом элементов одного и того же порядка и соответствующую разным формам «обычной» физической энергии, и «радиальную», «которая влечет его в направлении все более сложного и внутренне сосредоточенного состояния». Радиальная энергия способствует возрастанию сложности и упорядоченности организации, т. е. является движущей силой прогрессивной эволюции.
Рациональным зерном в подобных концепциях является понимание закономерного (хотя и абсолютизируемого в метафизическом духе) характера эволюции по пути морфофизиологического прогресса. Этого понимания, к сожалению, недостает некоторым современным неодарвинистским концепциям, концентрирующим внимание на процессах микроэволюции и подчеркивающим «оппортунистический характер» отбора.
Как уже отмечалось, естественный отбор не несет в себе никаких предпосылок, которые направляли бы эволюцию именно по пути морфофизиологического прогресса. Это подчеркивал еще Ч.Дарвин, полагавший, что эволюционный прогресс является одним из следствий борьбы за существование: отбор способствует совершенствованию организмов по отношению к условиям их существования; при неуклонно нарастающем общем усложнении биотической среды (благодаря дивергенции видов) это ведет к постепенному общему повышению организации.
Развивая идею Ч.Дарвина, некоторые современные ученые, например Ю.М.Оленов, полагают, что усложнение биотической среды в качестве движущей силы прогрессивной эволюции выражается в усложнении структуры биоценозов и межвидовых взаимодействий. Усложнение биоценозов и биосферы в целом, несомненно, происходило в истории Земли, однако между этими процессами и арогенезом нет простых и однозначных соотношений. Общий характер организации экосистем (определяемый обилием видов, входящих в их состав, различными формами взаимодействий между ними, соотношениями процессов биосинтеза и деструкции органических веществ и путями передачи продуктов биосинтеза в биоценозе), вероятно, не претерпел принципиальных изменений по крайней мере с конца палеозоя, когда в основном была заселена вся суша и сложились наземные экосистемы (водные экосистемы сформировались еще раньше). Разумеется, происходили изменения видового состава биоценозов, во многих случаях — значительные, возникали новые виды и новые высшие таксоны растений и животных, изменялся уровень биопродукции и распада органических веществ, многие экосистемы распадались и формировались новые. Особенно крупные преобразования биоценозов (биоценотический кризис) произошли на рубеже мезозоя и кайнозоя. Однако нет серьезных оснований полагать, что современные экосистемы организованы принципиально иначе (не в частностях, а по общей структуре) и сложнее, чем мезозойские.
В то же время на протяжении мезозоя и кайнозоя в разных филогенетических стволах возникали ароморфозы, далеко продвинувшие эволюцию по пути морфофизиологического прогресса (достаточно упомянуть формирование организации птиц и млекопитающих, появление человека). Нужно еще раз подчеркнуть, что эти ароморфозы отнюдь не были приурочены к каким-либо значительным изменениям условий внешней среды. Например, формирование организации птиц происходило во второй половине юрского периода, когда физико-географические условия не претерпели значительных перемен. С другой стороны, существенные преобразования биоценозов часто происходили уже после возникновения новых групп (например, во второй половине мезозоя в процессе расселения по Земле покрытосеменных растений).
Таким образом, арогенез не обнаруживает отчетливой зависимости от преобразований экосистем. К морфофизиологическому прогрессу, как и к другим проявлениям направленности эволюции, в полной мере можно применить уже упоминавшееся мнение Ч.Дарвина, что характер преобразований организмов определяется в гораздо большей мере природой их организации, чем характером изменений внешних условий (хотя, как мы видели, сам Дарвин не связывал это положение с эволюционным прогрессом). Логично заключить, что специфика орогенеза в значительной степени определяется свойствами самих организмов.
В то же время все ароморфозы имеют отчетливо приспособительный характер, будучи связаны с теми или иными конкретными адаптациями — совершенно различными у разных групп организмов. Следовательно, морфофизиологический прогресс, как и прочие направления эволюции, идет под контролем естественного отбора. Но при этом, в отличие от других направлений эволюции, арогенез имеет поступательный характер: последовательные ароморфозы, связанные с разными адаптациями, интегрируются, что выражается в общем повышении уровня организации.
Г. Стеббинс сделал попытку объяснить поступательный характер эволюционного прогресса в рамках редукционистской концепции «оппортунистического» действия естественного отбора. Он предположил, что повышение уровня организации является побочным результатом развития некоторых «оппортунистических» (т. е. возникающих ненаправленно) адаптации, достигнутый же уровень организации сохраняется — вплоть до нового случайного его повышения — просто благодаря приспособительному значению соответствующих характеристик организма.
Однако из допущения принципиальной равнозначности и ненаправленности возникновения любых адаптации следует, что равно вероятно возникновение новых адаптации, как повышающих, так и понижающих общий уровень организации, и их эффект в отношении уровня организации должен уравновешиваться. С другой стороны, при изменениях внешних условий приспо-собительное значение признаков организма неизбежно значительно изменяется, поэтому представления о сохранении достигнутого уровня организации как следствии приспособительного значения его характеристик являются упрощенными и неточными. Наконец, понимание эволюционного прогресса как случайного побочного результата равнозначных «оппортунистических» адаптации игнорирует реальную разнокачественность приспособлений, и в частности специфику ароморфозов и эпектоморфо-зов как изменений, имеющих общее значение для организма в целом. Иными словами, концепция Стеббинса полностью игнорирует эволюционную роль системной организации целостных организмов.
Для выяснения причин поступательного характера морфофи-зиологического прогресса необходимо выяснить специфику ароморфных адаптаций по сравнению с наиболее обычными, т. е. алломорфными.
Чтобы интегрироваться естественным отбором в поступательном процессе прогрессивной эволюции, разные ароморфозы, связанные с различными адаптациями, должны, во-первых, иметь общие черты и, во-вторых, обеспечивать важные преимущества в борьбе за существование. Общими для любых ароморфозов являются именно те основные критерии морфофизиологического прогресса, которые мы рассматривали в начале этой главы. А особые преимущества ароморфных изменений определяются важнейшими общими особенностями организации и функционирования живых систем.
Для существования организмов в изменчивой внешней среде необходимым условием является их специфическое обособление от последней при контролируемом живыми системами обмене веществ и энергии, обеспечивающем сохранение динамического равновесия организмов. Эволюционные изменения, соответствующие главным критериям морфофизиологического прогресса (дифференциация, интеграция, рационализация и др.) способствуют выполнению этого основного условия существования живых систем, повышая независимость внутренней среды организма от внешних воздействий. Тем самым ароморфозы повышают общую устойчивость живых существ в изменчивых условиях. Поэтому они имеют универсальное значение для разных групп организмов, что и определяет широкое распространение и поступательный характер арогенеза в разных филогенетических стволах.
Хотя морфофизиологический прогресс в наибольшей степени характерен для групп организмов, образ жизни которых требовал повышения активности основных биологических функций (питание, газообмен, передвижение, размножение), даже в филогенезе автотрофных прикрепленных растений происходили несомненные ароморфные изменения. Таковы дифференцировка вегетативного тела у высших растений на различные ткани и органы, развитие проводящей, покровной и механической систем, возникновение семени и т. п. Эти преобразования соответствуют по крайней мере первым пяти основным критериям арогенеза. Проявления морфофизиологического прогресса в эволюции неподвижно прикрепленных растений не противоречат сказанному выше о регрессивных изменениях у прикрепленных животных. Катагенез наблюдается при переходе от активного подвижного образа жизни к прикрепленному, тогда как при постоянно прикрепленном образе жизни проявляется действие общей закономерности эволюции — развития в направлении арогенеза, хотя и значительно слабее выраженного, чем у активно подвижных организмов.
Специфические приспособления, связанные с конкретной средой обитания и образом жизни данной группы, нередко накладывают разнообразные морфофизиологические ограничения и запреты на дальнейшее развитие в направлении арогенеза. Для высших наземных растений автотрофность определяет относительно невысокую общую интенсивность обмена веществ и энергии; необходимость извлечения из почвы воды и минеральных солей требует развития корневой системы, которая, подобно якорю, прочно удерживает растение на месте; прикрепленный образ жизни делает ненужным развитие приспособлений для передвижения, органов чувств и общего координационного центра (нервной системы). Более того, естественный отбор благоприятствует сохранению относительно слабой интеграции растительного организма, поскольку с ней связана более высокая способность к регенерации (восстановлению утраченных частей тела) и к вегетативному размножению, чрезвычайно важная для растений, поскольку они не могут активно защищаться или избегать растительноядных животных и разного рода механических повреждений.
Точно так же во многих филогенетических стволах животных специфические адаптации могут сдерживать или препятствовать дальнейшему общему морфофизиологическому прогрессу. Например, сдерживает арогенез значительное развитие органов пассивной защиты, подобных раковине моллюсков, панцирю черепах и т. п. Однако эти ограничения не являются абсолютными: у голо-воногих моллюсков раковина была преобразована в гидростатический аппарат, а затем редуцировалась, и современные голово-ногие, как известно, достигли значительной высоты организации, вероятно не уступающей по сложности и совершенству «среднему уровню» позвоночных.
Выше мы уже рассматривали комплекс морфофизиологичес-ких ограничений и запретов в организации земноводных, связанных каскадом корреляций с ключевым запретом — несовершенным механизмом вентиляции легких. У предков высших позвоночных — амниот — развился эффективный механизм вентиляции легких — всасывающий дыхательный насос грудной клетки. Развитие этого ароморфоза повлекло за собой каскад важных перестроек организации, который можно схематизированно представить следующим образом: 1) исчезла необходимость кожного дыхания; 2) в покровах усилились процессы ороговения эпидермиса, и это позволило ликвидировать непрерывные потери влаги в воздушной среде за счет испарения с поверхности влажной кожи и обеспечило лучшую механическую защиту; 3) одновременно открылись возможности для последующей дифференциации производных покровов (развитие роговых чешуи рептилии, перьев птиц, волос млекопитающих); 4) ороговение эпидермиса позволило также решить проблему осморегуляции в водной среде, защитив организм от обезвоживания в морской воде и от переобводнения в пресной; 5) в связи с этим появилась возможность сформировать в выделительной системе физиологический механизм экономии воды за счет ее обратного всасывания из мочи и синтеза концентрированных продуктов белкового обмена, все эти преобразования открыли доступ амниотам в широкий круг наземных местообитаний, включая самые засушливые, а также в море; 6) исчезла необходимость смешивать в желудочке сердца кровь от легких и кожи, в результате появилась возможность для разделения артериальной и венозной крови в сердце внутрижелудочко-вой перегородкой, что, в свою очередь, привело к перестройкам отходящих от сердца артериальных стволов и открыло возможность повышения уровня метаболизма и развития гомойотермии; 7) утрата ротоглоточной полостью и подъязычным аппаратом роли дыхательного насоса позволила развить механически более совершенные конструкции черепа (у земноводных череп должен быть широким, низким и плоским для эффективной работы подъязычного дыхательного насоса).
Ароморфозы, существенно изменяющие взаимодействия различных систем организма, снимая существовавшие прежде эволюционные запреты и ограничения на пути арогенеза и открывая новые возможности функционирования и эволюционных преобразований всего каскада зависимых систем, были названы ключевыми. Ключевые ароморфозы открывают путь другим важным изменениям организации, которые сами также могут быть ароморфозами, но не могут развиться без предварительного осуществления ключевого ароморфоза.
Ключевые ароморфозы возникали на всех этапах развития жизни. Так, важнейшим ключевым ароморфозом было приобретение предками эукариот в середине протерозоя актино-миозиновой сократительной системы, обеспечивающей цитоплазматическую подвижность. Ее появление могло быть связано с хищническим образом жизни и развитием фагоцитоза у предков эукариот. Развитие подвижности цитоплазмы потребовало обособления ядерных структур от остальной цитоплазмы, с этим связано возникновение ядерной оболочки из специализированной части цитоплазматического ретикулума. Актино-миозиновая система внутриклеточной подвижности позволила решить проблему клеточного выделения и осморегуляции — сформировался аппарат пульсирующих вакуолей, функция которых основана на использовании этих же сократительных белков. Сократительные белки функционируют и в аппарате микротрубочек, формирование которого было, вероятно, первоначально связано с обслуживанием органелл клеточного передвижения — жгутиков. Только после развития актино-миозиновой системы стало возможно возникновение упорядоченного механизма клеточного деления — митоза.
Ключевые ароморфозы (как и другие ключевые изменения организации) оказывают глубокое воздействие на направления и темпы макроэволюционных преобразований; эти вопросы будут рассмотрены в следующей главе.
Д.Кэмпбелл обратил внимание еще на один важный аспект эволюционного прогресса: по мере совершенствования организации биологические системы приобретают возрастающую способность контролировать собственную эволюцию. Это выражается, в частности, в приобретении клетками специальных комплексов энзимов (ДНК-полимераза, ДНК-лигаза, репликаза, мутаза, ре-комбиназа, транслоказа, инвертаза и др.), которые катализируют самые различные изменения молекулы ДНК, определяя возникновение соответствующих мутаций. Согласно Кэмпбеллу, гены, эволюционировавшие в определенном направлении, часто связаны в геноме с особым механизмом, повышающим способность мутировать именно в этом направлении.
В связи с этим важно отметить, что высшие организмы эволюционируют иными и притом более экономичными способами, чем более примитивные. Если основные приспособительные эволюционные изменения у прокариот связаны с глубокими биохимическими преобразованиями и происходят при выбраковке отбором огромного числа особей в каждом поколении, то у эукариот — с морфологическими преобразованиями при относительно незначительных биохимических изменениях, и только для высших животных характерны приспособительные изменения поведения, позволяющие в определенной степени снизить необходимость морфологических перестроек. Очевидно, в ходе арогенеза изменяется сам характер эволюционных преобразований.
Вероятно, повышение возможностей контроля со стороны организмов за своими эволюционными преобразованиями и становление все более гибких форм «эволюционного реагирования» организмов на изменения внешней среды можно рассматривать как еще один важный показатель арогенеза. Однако заметные сдвиги по этому показателю можно обнаружить лишь в самой широкой исторической перспективе, сравнивая качественно различные уровни организации.
Сопоставив все критерии и проявления морфофизиологиче-ского прогресса, можно заключить, что общим итогом и сущностью арогенеза является повышение независимости организма от внешней среды. Морфофизиологический эволюционный прогресс является закономерностью эволюции, обусловленной важнейшими общими свойствами живых систем и прежде всего целостных организмов: необходимостью поддерживать системный гомеостаз при обмене веществ и энергии с внешней средой. Таким образом, закономерное совершенствование организации живых систем в эволюции является следствием специфических особенностей самой этой организации.
Однако при этом необходимо учитывать и важнейшую роль естественного отбора. Как мы видели, ароморфозы развиваются на основе определенных приспособлений и сами являются особого рода приспособлениями, повышающими независимость организма от внешних условий. Единственным же фактором, обусловливающим развитие любых приспособлений, является естественный отбор. Очевидно, отбор представляет собой движущую силу арогенеза, без участия которой невозможна реализация никаких тенденций эволюционного развития, определяемых особенностями системной организации живых существ.
Среди специфических черт макроэволюции пульсирующий характер ее темпов, выражающийся в виде чередования периодов типогенеза и типостаза, более всего привлекал внимание исследователей. Именно эта закономерность макрофилогенеза способствовала возникновению сальтационных гипотез. Как уже отмечалось, типогенез имеет характер квантовой эволюции, при которой значительные преобразования организации в процессе формирования основного комплекса признаков нового крупного таксона происходят в течение относительно краткого промежутка времени. Напомним, что о краткости сроков здесь говорится в масштабах макрофилогенеза, для которого относительно краткими можно считать промежутки времени в 10—30 млн лет по сравнению с сотнями миллионов лет периодов типостаза. Выше (гл. 2 этой части) мы рассмотрели ряд гипотез, предложенных в рамках синтетической теории эволюции для объяснения высоких темпов типогенеза (смена адаптивных зон, «генетические революции», видовой отбор, биоценотическая регуляция и преобразования экосистем). Эти гипотезы, (особенно первая и последняя из них) вносят определенный вклад в решение данной проблемы, но каждая из них имеет отношение лишь к некоторым частным случаям.
Вероятно, квантовый эффект в макрофилогенезе является внешним выражением действия нескольких эволюционных механизмов, основанных на системных свойствах организмов и объединяемых пороговым характером своего проявления.
Самый простой вариант такого механизма представляют эволюционные перестройки функциональных (эргонтических) корреляционных систем фенотипа. В целостной корреляционной системе изменение одного компонента требует соответствующих изменений других компонентов. Например, невозможно изменить форму суставной поверхности на одной кости без соответствующих изменений другой кости, сочлененной с первой (см. также примеры динамических координации, с. 366). В сущности, коррелятивные связи в морфофункциональных системах организма сами являются причиной возникновения сил мощного направленного отбора, требующих гармонических перестроек целого по принципу «все или ничего».
При этом очевидно, что одновременное или строго последовательное возникновение всех необходимых мутаций разных структурных генов, которые могли бы обеспечить гармоничное преобразование всех затронутых компонентов фенотипа, чрезвычайно маловероятно. Поэтому на первых этапах эволюционных преобразований таких корреляционных и координационных систем важную роль могут играть модификационные (ненаследственные) изменения взаимосвязанных элементов (рис. 109). Хотя такие изменения остаются в пределах прежней нормы реакции, они в совокупности могут привести к существенным преобразованиям общего фенотипического облика в очень короткие филогенетические сроки. Для палеонтолога, изучающего ископаемые остатки организмов, такие преобразования будут выглядеть как макроэволюционные.
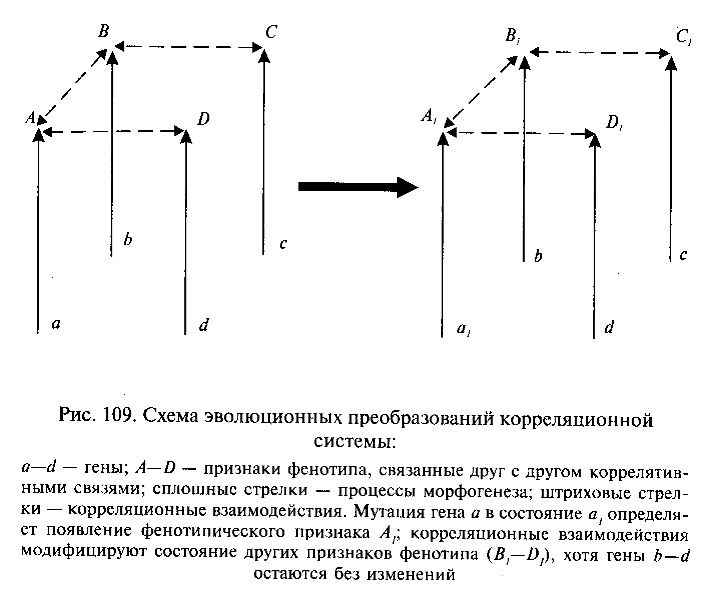
Проиллюстрировать действие этого механизма проще всего на примере модификаций, компенсирующих различные повреждения организма, возникающие в естественных условиях или в эксперименте. Г. Гюнтер и Д. Уорд обнаружили среди отловленных в море окунеобразных рыб, принадлежащих к разным видам (Chaetodipterus faber, Micropogon undulatus и др.), несколько особей, у которых был полностью утрачен хвостовой плавник (вероятно, в результате нападения хищника), причем отсутствие хвоста было компенсировано разрастанием назад спинного и анального плавников и их мускулатуры. Эти рыбы были вполне жизнеспособны и, очевидно, прожили уже значительное время после утраты хвостового плавника. Конечно, в данном случае произошло не наследственное изменение организации, а лишь комплекс модификационных ненаследственных изменений, компенсирующих повреждение. Однако можно полагать, что если утрата хвостового плавника будет обусловлена мутацией, то организм сможет компенсировать ее неблагоприятные последствия посредством аналогичных модификационных перестроек.
Интересные модификации скелетно-мышечной системы возникают у четвероногих млекопитающих в ответ на экспериментальное приучение к постоянному передвижению на задних конечностях. У обезьян-макаков это приводило к возникновению характерного для человека S-образного изгиба позвоночника; у крыс — к некоторому укорочению (брахицефализации) черепа, а также к усилению ягодичных мышц и изменениям формы таза, связанным с увеличением угла между лобковой и подвздошной костями. Особенно любопытно, что эти модификации в определенной степени сходны с изменениями соответствующих структур в эволюции предков человека, несомненно, связанными с переходом к двуногому передвижению.
В этом отношении интересны также результаты экспериментов И.Гармса с илистыми прыгунами (Periophthalmus) — окунеобразными рыбами (рис. 110), обитающими в тропической приливо-отливной мангровой зоне. Во время отливов илистые прыгуны регулярно остаются на суше, где они могут переползать и даже совершать прыжки, отталкиваясь передними плавниками. Гармс выдерживал илистых прыгунов длительное время на суше и подвергал их воздействию повышенной концентрации тироксина (гормон щитовидной железы). В результате у рыбок уплотнялись ткани внутренних органов и кожа, усиливалось ороговение эпидермиса, удлинялись конечности, значительно уменьшались отолиты (слуховые камешки во внутреннем ухе, которые у рыб обычно имеют очень крупные размеры). Эти адаптивные модификации сходны с соответствующими изменениями, происходящими в онтогенезе земноводных при метаморфозе, который стимулируется повышенной секрецией тироксина щитовидной железой. Можно полагать, Что эти изменения, приспосабливающие организм земноводных к жизни на суше, также могли первоначально представлять собой модификации, возникавшие в ответ на повышенную тироксинизацию, а эта последняя бьыа результатом мутации, обусловившей усиление функции щитовидной железы.

Если возникшие подобным образом изменения фенотипа имеют адаптивную ценность, стабилизирующий отбор будет благоприятствовать наследственному закреплению новой комбинации фенотипических признаков и автономизации их онтогенетического развития. При этом отбор способствует закреплению и распространению мутаций, обеспечивающих формирование этих признаков в онтогенезе без модифицирующего влияния внешних условий — т. е. генокопий соответствующих модификаций. В связи с этим напомним подмеченный Р. Гольдшмидтом параллелизм фенотипического проявления мутаций и модификаций (см. с. 61). Дальнейшие перестройки онтогенеза в направлении автономизации развития этих признаков приведут ко все более ранней их закладке в эмбриогенезе. Отбор также будет способствовать новому расширению нормы реакции в том же направлении для создания нового «запаса прочности», нового резерва модификационной изменчивости, который может быть использован в последующей эволюции.
Таким образом, целостные системы организма, связанные функциональными корреляциями, преобразуются в эволюции как целое. Поэтому, как показал Л.Ридл, их преобразования должны происходить гораздо быстрее, чем при отборе независимых случайных вариаций отдельных признаков.
Важное значение для возникновения высоких темпов типогенеза имеют каскадные взаимодействия между разными системами организма, рассмотренные в двух предшествовавших главах. Ключевые изменения являются своего рода «спусковым механизмом», запускающим весь каскад эволюционных преобразований в зависимых системах, имеющих важное адаптивное значение (как мы уже отмечали, эти преобразования бьыи и раньше возможны на генетическом и морфогенетическом уровнях, но запрещены на морфофизиологическом, см. с. 365). После приобретения ключевого изменения при сильном давлении отбора, благоприятствующего развитию этих важных адаптации, эволюционные изменения должны происходить с максимально возможной скоростью. Таким образом, ключевые изменения организации являются своего рода «эволюционным триггером», значительно увеличивающим темпы макроэволюционных преобразований.
Происходящие при этом микроэволюционные изменения ничем не отличаются от обычного видообразования, и темпы микроэволюции в периоды каскадных макроэволюционных преобразований могут оставаться на среднем уровне. Однако взаимозависимость перестроек разных систем организма, скоординированных и разрешенных ключевым преобразованием, сама становится источником для возникновения мощного давления направленного отбора, благоприятствующего изменениям в заданном направлении. Скоординированность и направленность преобразований системной организации значительно ускоряют макроэволюцию.
Одним из наиболее известных примеров является каскад эволюционных преобразований у предков рептилий, связанный с развитием у них ключевого ароморфоза — механизма дыхательных движений грудной клетки, ставшего высокоэффективным всасывающим легочным насосом. Как мы видели в предыдущей главе, развитие этого насоса ликвидировало весь комплекс морфофизиологических ограничений и запретов, существующих в организации амфибий и обусловленных недостаточностью примитивного нагнетательного легочного насоса (подъязычного аппарата) для выведения из легких углекислого газа.
Возникновение направленности и в силу этого — высоких темпов макроэволюционных преобразований может быть обусловлено не только каскадными взаимодействиями, но и другими формами координации между различными системами организма — без ясно выраженного доминирования одной системы над другими (хотя квантовый эффект при этом, очевидно, будет выражен слабее). Такой характер макроэволюции соответствует обоснованной Т. Кемпом модели взаимосвязанного прогресса, согласно которой формирование организации нового крупного таксона происходит путем серий взаимосвязанных изменений различных систем организма. По этой модели происходило развитие основных признаков организации млекопитающих в филогенезе высших синапсидных рептилий — териодонтов. При этом отсутствие общего межсистемного ключевого признака не исключает каскадных взаимодействий в отдельных системах.
Так, сложный комплекс преобразований челюстного аппарата у териодонтов был обусловлен ключевой адаптацией — совершенствованием переработки пищи челюстями. Эти преобразования включают дифференциацию зубов на резцы, клыки и трехбугорчатые щечные зубы («коренные»), увеличение размеров зубной кости нижней челюсти и ее венечного отростка, на который прикреплялась височная мышца (роль последней усиливалась при развитии механизма пережевывания пищи). Развитие этих признаков непосредственно повлекло за собой изменение общих пропорций черепа (увеличение объема височных ям, вмещающих жевательную мускулатуру, привело к относительному увеличению заглазничной части черепа), развитие вторичного челюстного сустава, разрастание зубной кости и редукцию остальных костей нижней челюсти и преобразование костей первичного челюстного сустава в косточки среднего уха. Пережевывание пищи требует жесткого давления на перерабатываемый корм, что привело к утрате примитивной подвижности нёба и верхней челюсти (кинетизма черепа). С другой стороны, задержка пищи для переработки в ротовой полости приводит к задержке дыхания, крайне нежелательной при развитии гомойо-термии. Это вызвало необходимость развития вторичного нёба, разделившего носовой ход и собственно ротовую полость. Кроме того, вторичное нёбо укрепляет верхнюю челюсть, что также важно при пережевывании пищи. Развитие всех этих преобразований было обусловлено указанной ключевой адаптацией. Весь комплекс изменений развивался скоррелированно и направленно, что доказывается многочисленными параллелизмами в разных филетических линиях териодонтов. Это определило высокие темпы типогенеза.
Ускорению темпов типогенеза могут способствовать и другие эволюционные механизмы, обладающие пороговым характером проявления. Например, таковым является механизм морфофункциональной преадаптации, посредством которой в филогенезе происходит смена адаптивной роли органов. После прохождения стадии преадаптационного порога перед данной группой организмов открываются новые адаптивные возможности, которые реализуются в относительно краткие (в масштабах макроэволюции) сроки. Одновременно происходит перестройка всего комплекса структур, связанных корреляциями с органом, приобретшим новую адаптивную роль.
Возникновению квантового макроэволюционного эффекта могут также способствовать дискретные изменения программ онтогенеза. Взаимодействие различных факторов в морфогенезе часто имеет пороговый характер. В этом случае сравнительно незначительные генетические перестройки, обусловленные мутациями генов-регуляторов (особенно типа гомеобоксных генов, контролирующих экспрессию целых генетических систем) могут привести к существенным преобразованиям фенотипических признаков. Например, при обработке эмбриональных зачатков кожи ретиноевой кислотой (витамин А) зачатки роговых чешуи на конечностях у эмбрионов птиц преобразуются в перья, а зачатки волос у эмбрионов млекопитающих — в железистые структуры.
В эволюции корреляционных систем изменения их концевых звеньев и боковых ветвей могут происходить более или менее независимо друг от друга — вплоть до соответствия принципу независимой эволюции разных признаков по Г. Осборну. Скорость этих изменений определяется только факторами микроэволюции, в первую очередь частотой мутаций и давлением отбора. Такие изменения происходят наиболее часто и являются основой для процессов видообразования и адаптивной радиации, не затрагивающих основной план строения крупного таксона.
Гораздо реже возникают изменения на более глубоких уровнях организации корреляционных систем. Их следствием являются координированные изменения всех взаимосвязанных компонентов. В результате происходят дискретные преобразования целого комплекса признаков, в том числе в общем плане строения. Эти преобразования также осуществляются через обычные микроэволюционные механизмы, но их координированный и направленный характер обусловливает высокие темпы макроэво-люционных перестроек. Р.Ридл показал, что темпы эволюционных преобразований интегрированных целостных систем могут быть на несколько порядков выше, чем эволюция, основанная на комбинировании независимых вариаций отдельных компонентов.
Очевидно, при необходимости быстрых координированных перестроек гетерогенного адаптивного комплекса отбор должен благоприятствовать возникновению корреляционных связей между разными его компонентами и подчинению разных признаков общему контролю в онтогенезе, вплоть до формирования целостных генетических комплексов. При этом и на молекулярном уровне формируется блочно-иерархическая организация генома, позволяющая существенно увеличить скорость эволюции за счет действия отбора на целостные блоки в геноме, контролируемые общими генами — операторами и регуляторами (см. с. 90). Иначе говоря, длительно действующий направленный отбор может благоприятствовать более глубокой интеграции систем организма, которая способствует значительному убыстрению макроэволюции, поскольку эволюционные преобразования целостной системы происходят несравненно быстрее, чем комбинирование благоприятных мутаций по отдельным признакам.
Таким образом, можно заключить, что высокие темпы типогенеза определяются прежде всего самим характером происходящих изменений организации, причем осуществляется своего рода самоконтроль темпов макроэволюции со стороны организма.
Все формы корреляционных и координационных связей в целостном организме при возникновении изменений в ключевых системах способствуют комплексному характеру макроэволюционных преобразований, что многократно повышает темпы типогенеза. С другой стороны, стабильность общего плана строения в период типостаза обеспечивается разными механизмами системного гомеостаза, действующими на всех уровнях организации, — канализация морфогенеза под воздействием генов-модификаторов, регуляция возникающих изменений при взаимодействиях разных морфогенетических систем, мультифункциональность и функциональная пластичность фенотипических структур и др. (Мультифункциональность — выполнение данным органом или структурой нескольких функции, связанных с разными адаптациями. В этом смысле термин «мультифункциональность» иногда заменяют на «мультиадаптивность»)
Вообще степень эволюционного консерватизма данной системы в целом пропорциональна ее способности к авторегуляции. Например, в скелетно-мышечном комплексе мускулы обладают более высокой онтогенетической пластичностью, чем кости; в онтогенезе строение и функции мышц относительно легко модифицируются в соответствии с изменениями нагрузок. К тому же мышцы обладают высокой функциональной лабильностью, т. е. способны обеспечивать различные оттенки движений сокращением разных пучков мышечных волокон в различных комбинациях. Иными словами, мышцы обладают высокими авторегуляционными возможностями. Следствием этого является возможность для данного мышечного комплекса удовлетворительно функционировать при разнообразных адаптивных требованиях, соответствующих различным направлениям отбора. Скелетные же элементы обладают авторегуляционными способностями в значительно меньшей мере, чем мышцы. Поэтому они не могут адекватно реагировать адаптивными модификациями на существенные изменения условий функционирования и должны отвечать на изменения направления отбора наследственными перестройками, основанными на соответствующих мутациях. В результате этого в эволюции скелетные компоненты в целом оказываются менее консервативными, чем мышечные.
При этом целостные морфофункциональные системы (например, скелетно-мышечные комплексы конечностей или челюстного аппарата) проявляют более высокий эволюционный консерватизм, чем отдельные их компоненты (в данном примере кости и мышцы). Это связано с более значительными возможностями авторегуляции систем высокого ранга, в которых авторегуляционные приспособления отдельных элементов дополняются соответствующими свойствами всей системы. Еще Галену было известно, что целый череп прочнее по отношению к ударам, чем его отдельные кости. Причиной этого являются амортизационные свойства шовных соединений между костями.
У многоклеточных организмов авторегуляционные возможности основных морфогенетических систем, обеспечивающих в онтогенезе формирование общего плана строения данной группы, в целом значительно выше, чем таковые генотипа или отдельных фенотипических признаков. Поэтому, как уже бьшо отмечено в главе об эволюции онтогенеза, аппарат наследственности преобразуется в макрофилогенезе значительно быстрее, чем основные фенотипические структуры. Наиболее устойчивы и консервативны в макроэволюции эпигенетические корреляционные системы организма, обладающие наибольшими авторегуляционными возможностями.
Таким образом, корреляционные системы организма, представляющие собой гомеостаты, проявляют тенденцию к эволюционной стабильности. Можно допустить, что нормальное функционирование таких систем происходит в определенных режимах, действующих в пределах основных состояний или типов их конструкции, промежуточные состояния между которыми функционально неэффективны и поэтому эволюционно неустойчивы. Это обусловливает дискретность макроэволюционных преобразований, т. е. их квантовый характер. Макроэволюция представляет собой переход от одного устойчивого состояния морфофункциональных, морфогенетических и генетических систем организмов к другому их устойчивому состоянию.
В целом можно сказать, что характерные пульсирующие темпы макроэволюции, т. е. чередование типогенеза и типостаза в макрофилогенезе, являются следствием основных свойств корреляционных биологических систем, и в первую очередь — целостного организма.