
Проблемы Эволюции |
Экосистемные перестройки и эволюция биосферы. Москва, "Недра", 1994. С. 25-32.
Едва ли не самым значительным событием в истории биосферы было возникновение эвкариот. Это событие послужило началом кардинальной перестройки глобальной экосистемы. Существовавшие в архее и раннем протерозое сети биогеохимических взаимодействий микробных сообществ стали надстраиваться трофической пирамидой эвкариот. Эта, аддитивная, фаза эволюции глобальной экосистемы характеризовалась формированием все новых трофических уровней, наивысшим из которых стал уровень многоклеточных животных. Дальнейшее существование морских экосистем можно представить как процесс непрерывного замещения отдельных членов трофической иерархии при том, что структура в целом оставалсь стабильной [I].
Появление эвкариот и усложнение трофической структуры глобальной экосистемы можно рассматривать как неизбежный автогенетический процесс. Но возникает вопрос, почему и когда это было разрешено?
Толерантность бактерий к различным условиям среды обитания, основанная на широком спектре механизмов бактериальной адаптации, служит и служила залогом поразительной устойчивости прокариотной экосистемы, нс с другой стороны, -и причиной ее консервативности [10]. Важнейшие механизмы бактериальной адаптации, прежде всего - быстрые мутации в направлении новых циклов метаболизма, латеральная передача "полезных" генов, уменьшение скоростей метаболизма до крайне низких -уровней, что может сопровождаться спорообразованием, высыханием, миниатюризацией, а может протекать и без морфологических изменений, - все это ставит бактериальные сообщества в такое положение, что скорее весь мир многократно изменится, чем они.
Судя по данным палеонтологической летописи архея и протерозоя, цианобактериальные сообщества (преимущественно сохранившиеся в ископаемом состоянии ввиду более крупных размеров цианобактерий, чем других бактерий), прошли через всю историю Земли без существенных морфологических, а, возможно, и биохимических, изменений. Они явно доминировали во всех мес-тообитаниях. Что же заставило их потесниться?
Можно предположить, что невероятная толерантность бактерий к разнообразным физико-химическим факторам среды отражает условия, существовавшие на ранней Земле в архее, коща и сформировались основные механизмы бактериальной адаптации. И если это справедливо, то верно и то, что многие из этих параметров, оптимальные для роста биомассы бактерий, могли необратимо уйти от оптимума еще в докембрии.
Действительно, ранние этапы истории нашей планеты были связаны с высокими температурами, химически разнообразными, в том числе и агрессивными, средами, в которых могли существовать только прокариоты.
Что же изменилось в среде обитания на протяжении докембрийского зона? Избегая количественных характеристик, зачастую противоречивых, отметим лишь общие тенденции в изменении физико-химических параметров биосферы, которые могли негативно влиять на прокариотные сообщества.
Во-первых, постепенно уменьшался объем геохимических ресурсов энергии, пригодных для прокариот. Об этом можно судить по снижению вулканической и гидротермальной активности на протяжении протерозоя [9] также и потому, что огромные массы жизненно важных химических элементов и их соединений были изъяты из биогеохимического круговорота и захоронены в виде рудных залежей осадочного происхождения.
Во-вторых, в значительной части обширных местообитаний средняя температура постепенно снижалась на протяжении до-кембрия, достигнув значений ниже оптимума многих групп прокариот. Об этом могут свидетельствовать следы крупных оледенении, древнейшее из которых (Гуронское) имело место около 2,3-2,5 млрд лет назад. Начиная приблизительно с 850 млн лет назад, ледниковые периоды становятся обычным и регулярным явлением геологической истории [5].
Не последнюю роль в глобальном похолодании до уровня балансирования вокруг точки нуля сыграли живые организмы, но ключевое значение имело, по-видимому, формирование обширных континентальных плит. С этим процессом связано увеличение площади мелководных местообитаний и возникновение апвел-линга. Оба эти явления способствовали более интенсивному круговороту важнейших метаболитов в морских экосистемах, что вело к росту первичной биопродуктивности. Рост биопродуктивности фотосинтезирующих организмов, в особенности цианобактерий, неизбежно сопровождался изъятием огромных масс углекислоты из воды и атмосферы, интенсификацией биогенного карбонатоосаждения и формированием карбонатных платформ [6, 7]. Снижение парникового эффекта происходило не только благодаря изъятию и захоронению углекислоты, но и благодаря окислению другого парникового газа - метана.
Еще одним фактором, подавлявшим развитие большей части прокариот, мог стать рост содержания свободного кислорода. После того, как основные массы биогенного кислорода перестали уходить на окисление металлов, и прежде всего - железа, этот ядовитый для многих групп прокариот газ стал накапливаться в гидросфере и атмосфере. Надо иметь в виду, что пороговые значения концентрации кислорода различны для разных групп прокариот. Самым заметным свидетельством микробной деятельности в докембрии принято считать строматолиты. Действительно, строматолитовые карбонаты являются важной составляющей осадочного чехла древних платформ. Мощности строматолитовых толщ протерозоя очень велики, и нужно иметь в виду крайне низкую скорость карбонатоосаждения современных аналогов строматолитов.
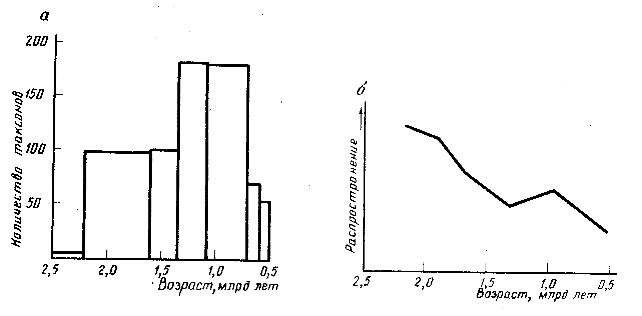
Рис. 1. Таксономическое разнообразие (а, по данным [18]) и распространение (б, по данным [7]) строматолитов в протерозое, определяемое по отношению объема строматолитовых пород на единицу мощности осадков на карбонатных платформах. Высокий вклад стромалитовых микробных сообществ в формирование карбонатных платформ в раннем протерозое был связан с более высокой карбонатной насыщенностью морских вод по сравнению с фанерозоем и с отсутствием конкурирующих высших водорослей, которые появились в позднем протерозое [3]
Небезынтересна динамика таксономического разнообразия этих биолитов, представляющих собой по сути побочный продукт функционирования микробных сообществ. Мы пока не представляем себе, что конкретно отражает морфология строматолитов -состав бактериальных сообществ или физико-химические параметры среды. Но факт, что разнообразие и обилие строматолитов растет на протяжении позднего протерозоя, достигая пика в районе 1 млрд лет назад, а затем довольно быстро падает (рис. 1, а, б).
Причины упадка строматолитовых сообществ активно обсуждаются. В числе основных гипотез (их критический обзор см. [7]) рассматриваются следующие: 1) появление эвкариотных водорослей, конкурирующих с цианобактериями за местообитание и ресурсы питания; 2) негативное влияние возникших многоклеточных животных, выедавших цианобактериальные дерновинки и нарушавших стабильность субстрата, на котором они росли; 3) уменьшение карбонатного насыщения морских вод.
Нам кажется, что эти гипотезы не исключают друг друга, а дополняют. Более того, мы могли бы добавить еще два фактора. Об одном из них упомянуто выше - это рост содержания свободного кислорода в атмосфере, высокие концентрации которого могли подавлять не только анаэробов, но и фотосинтезирующих цианобактерий. Надо иметь в виду, что к продуцированию кислорода в позднем протерозое подключились и эвкариотные водоросли.
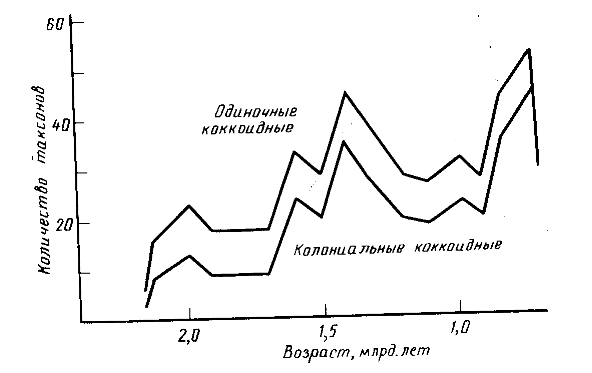
Рис. 2. Динамика видового разнообразия микрофоссилий из кремней в протерозое [15]
Еще один фактор - это длительная череда ледниковых периодов в промежутке от 850 млн лет до начала кембрия. И дело, вероятно, не столько в понижении средних температур, сколько в том, что в период роста ледниковых шапок уровень моря падал столь существенно, что обширные эпиплатформенные бассейны исчезали, и прежние бескрайние местообитания строматолитов сужались до узкой зоны окраины шельфа. Каждая постледниковая трансгрессия сопровождалась колонизацией мелководных местообитаний как бы заново, в обновленных условиях среды. И эти условия складывались не в пользу доминирующей роли прокариот.
Отражение всех этих событий можно видеть в кривых разнообразия ископаемых микроорганизмов - микрофоссилий из кремней, среди которых преобладают цианобактерий (рис. 2), и некоторых органикостенных микрофоссилий, часть которых также может быть отнесена к цианобактериям. Большие размеры и морфологическая сложность многих групп акритарх может свидетельствовать об их эвкариотной природе (рис. 3).
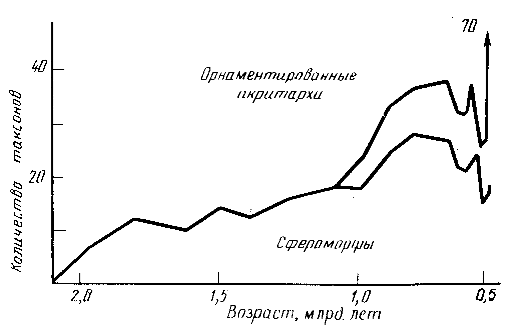
Рис. 3. Таксономическое разнообразие акритарх в протерозое [15]
На фоне растущего разнообразия микроорганизмов наблюдается и рост их размеров: формы диаметром 60-100 микрон известны уже 1850 млн лет назад, диаметром 200-600 микрон -1400 млн лет, более 1000 микрон - 1050 млн лет, относительно сложные акритархи появляются около 1000 млн лет, а крупные сфероморфы диаметром 1-7 мм были широко распространены около 850 млн лет назад. Все эти группы рассматриваются как вполне возможные остатки эвкариот, имея в виду, что верхний размерный предел прокариот находится в районе 60 микрон. Однако следует иметь в виду, что нижний размерный предел эвкариот -20-30 мкм.
Пик разнообразия эвкариотных организмов в докембрии приходится на интервал между 900-800 млн лет.
Между 850 млн лет и началом кембрия биосфера испытала серию перестроек, что выразилось в вымирании почти всех планктонных таксонов с размером более 600 микрон и многих относительно сложных "акритарх". Увеличение скорости захоронения органического углерода в эти кризисные периоды может свидетельствовать о соответствующем увеличении концентраций атмосферного кислорода. Но одновременно это может говорить и об олиготрофикации морских бассейнов. Косвенным подтверждением этого предположения является тенденция увеличения отношения поверхности по отношению к объему клетки у ископаемых микроорганизмов. Этот эффект достигается двумя способами: уменьшением размера клетки и появлением выростов, шипиков и других подобных приспособлений, увеличивающих поверхность обмена клетки с окружающей средой. Обе стратегии отмечаются в позднем рифее и в нижнем кембрии, хотя оли-готрофикация могла иметь разные причины.
Итак, если следовать данным анализа разнообразия строма-толитов и микрофоссилий с учетом изменения размерности последних, то можно думать, что эвкариоты появились на Земле не позднее 1,5 млрд лет назад. Косвенным подтверждением этому могут служить и две другие независимые линии свидетельств: данные филогенетической интерпретации молекулярной генетики [13, 14] и таксономическая интерпретация биомаркеров, в частности стеринов, извлекаемых из керогена или углеводородов протерозоя [16]. То есть предполагаемый момент появления эвкариот более чем на полмиллиарда лет предшествует максимуму количества и разнообразия строматолитовых сообществ и очень близок к моменту завершения крупнейшей эпохи накопления железистых кварцитов (примерно 1,7 млрд лет назад). Высказываются мнения и о более древнем происхождении эвкариот, основанные на интерпретации проблематичных ока-менелостей неясной природы [12].
Обычно происхождение эвкариот связывают с ростом содержания свободного кислорода в атмосфере до уровня, достаточного для аэробного дыхания.
Мы полагаем, что аэробное дыхание на прокариотном уровне должно было существовать задолго до появления аэробных эвкариот. Если стоять на позициях симбиогенеза эвкариот [2], то один из биотических компонентов, входивший в симбиоз, должен был обладать сформировавшимся механизмом аэробного обмена.
Вопрос этот может прозвучать и шире. Кто утилизовал в архее и раннем протерозое продукты жизнедеятельности бактериальных сообществ, и в частности биогенное органическое вещество? И если вопрос об аэробном дыхании в архее уже поднят [17], то на роль сапротрофов пока не найдено претендентов среди известных нам архейских и раннепротерозойских окаменелостей.
При этом упускается из виду, что локально высокие концентрации кислорода существовали в водных средах уже в раннем архее, когда содержание кислорода в атмосфере было крайне низким, а концентрации других газов, например COz, -на два порядка превышали нынешний уровень [9]. Высокие концентрации кислорода существовали в очень узкрй зоне - в поверхностном миллиметровом слое цианобактериальных матов, которые были широко распространены на протяжении большей части архея и особенно в протерозое.
Современные микробные маты могут рассматриваться как аналоги докембрийских строматолитов (особенно их пластовых разновидностей). Характерной особенностью бактериальных матов является резкий и флюктуирующий градиент кислорода и сероводорода. В течение светового дня уровень кислорода быстро поднимается внутри цианобактериальной фотической зоны (верхний миллиметр или чуть больше), а содержание сероводорода падает часто до нуля. В темное время суток фотосинтез угасает, и содержание кислорода падает практически до нуля, зато резко поднимается уровень содержания сероводорода [1, 11]
Подобная, весьма специфичная среда, чрезвычайно ограниченная по вертикали и весьма протяженная по горизонтали, и могла стать первичным биотопом для зарождающихся эвкариот.
Предлагаемая концепция первичного биотопа эвкариот в наибольшей степени соответствует теории симбиогенеза, в частности и потому, что вблизи поверхности цианобактериального мата соседствуют по меньшей мере три среды с различными геохимическими характеристиками и, следовательно, с различным составом микробных сообществ.
Именно эти три среды, а именно - уровень ниже фотической зоны мата, сама фотическая зона и узкая зона над поверхностью мата в водной среде, - могли стать источниками организмов, вошедших в симбиоз, который привел к формированию эвкариотной клетки.
Трудно надеяться, что мы сможем уловить момент появления эвкариот, тем более, что процесс симбиогенеза мог быть долгим, а в каких-то фазах - и обратимым. Он был приурочен к узкой зоне обитания. Значительно больше шансов распознать появившихся эвкариот в тот период, когда они начали играть существенную роль в экосистемах прошлого. Пока мы обратили внимание лишь на динамику количества и разнообразия строматолитов, предположив, что она может косвенно отражать и функционирование эвкариотной надстройки. В частности, падение количества и разнообразия строматолитов после 1 млрд лет назад можно определенно, связывать с возрастанием конкурентных отношений между прокариотными сообществами и эв-кариотами с явным выигрышем в пользу последних. Но до этого момента, по-видимому, имело место другое важное событие, а именно - выход эвкариот из пределов первичного биотопа в обогащенную кислородом водную среду.
1. Заварзин Г.А. Бактерии и состав атмосферы. - М.: Наука, 1984. -(Серия "Человек и окружающая среда").
2. Маргелис Л. Роль симбиоза в эволюции клетки. - М.: Мир, 1983.
3. Butterfield N.f., Knoll A.H. and Sweet K. Exeptional preservation of fossils an Upper Proterozoic shale // Nature. - 1988. - V. 334. -P. 424-427.
4. Christen R., Ratio A., Baroin A. Origin of mctazoans. A phytogeny deduced from sequences of the 28 S ribosomal RNA. In: Simonetta A.M. and S. Conway Morris (Eds.).
5. Chumakov N.M., Elston D.P. The paradox of Lale Proterozoic glaciations at low latitudes // Episodes. - 1989. - V. 12. - N2. -P. 115-120.
6. Fedonkin M.A. Neoproterozoic ecosystem restructuring: from net to pyramid // Walliser O.H. (Ed.): Phanerozoic global bio-events and event stratigraphy. Fifth Interbational conference on Bio-Events. Abstracts. Gottingen. - 1992. - P. 33-34.
7. Grotzinger f.P. Geochemical model for Proterozoic stromatolite decline // American Journal of Science. - 1990. - V. 290-A. - P.80-103.
8. Hasting J.F. Theoretical constraints on oxigen and carbon dioxide concentrations in the Precambrian atmosphere // Precambrian Research. V.34. - P. 205-229.
9. Hasting J.F. The Proterozoic atmosphere and climate // The Proterozoic Biosphere: A Multidisciplinary Study. A Symposium. University of California. Los Angeles. - 1988. - P. 55-56.
10. Knoll A.H., Bauld J. The evolution of ecological tolerance in procaryots. Transactions of the Royal Soc. of Edinburhg // Earth Sciences. - 1989. - N 80. - P.209-223.
11. Pierson B.K. Ecology and physiological characteristics of modern microbial mats: perspectives for interpretations of Proterozoic benthic communities // The Proterozoic Biosphere: a multidisciplinary study. A Symposium. University of Californaia, Los Angeles. - 1988. - P. 25-26.
12. Rozanov A. Yu. Evolution of Biosphere ai.d Skeletonisation 28th International Geological Congress // Abstracts. - Washington. D.C. 1989. - V.2. - P. 722-723.
13. Runnegar В. Molecular evidence for the history of Precambrian life // The Proterozoic Biosphere: a multidisciplinary study. A Symposium. University of California, Los Angeles. - 1988. - P. 24-25.
14. Schwartz R.M. and Dayhoff M.0. Origins of prokaryotes eukaryotes, mitochondria and chloroplasts // Science. - 1978. - N 199. - P. 395-403.
15. Sepkoski J.f. Patterns of diversity exhibited by the known Proterozoic fossil record // The Proterozoic Biosphere: a multidisciplinary study. A Symposium. University of California, Los Angeles. - 1988. -P. 57-58.
16. Summons R.E., Walter M.R. Molecular fossils and microfossils of procaryotes and protists from Proterozoic sediments // American Journal of Science. - 1990. - V.290-A. - P. 212-244.
17. Towe K.T. Aerobic respiration in the Archaean? // Nature. 1990. - V. 348. - N. 6296. - Р.54-56.
18. Walter M.R., Heys G.R. Links between the rise of the metazoa and the decline of stromatolites // Precambrian Research. - 1985. - V.29. -P. 149-174.
19. Woese G.R. Bacterial evolution // Microbiological Reviews. N 51. - P. 221-271.