
Проблемы Эволюции |
1968
Теория стабилизирующего отбора
III. ЭЛЕМЕНТАРНЫЕ ПРОЦЕССЫ ИЗМЕНЕНИЙ ОРГАНИЗМА В ЕГО ИСТОРИЧЕСКОМ РАЗВИТИИ
Так как прогрессивная эволюция организмов протекает в популяциях, то мы не могли оставить без рассмотрения основные процессы исторической изменяемости популяций. Они начинаются с неопределенных наследственных изменений (мутаций) отдельных особей и поддерживаются многократной повторностью этих изменений. Через процесс размножения разнообразные мутации распространяются под контролем естественного отбора во всей популяции. Постоянные скрещивания особей (панмиксия) ведут к созданию все новых комбинаций наследственных изменений. Естественный отбор непрерывно извлекает наиболее благоприятные в данных условиях, наиболее гармонические сочетания, а различные формы изоляции или вообще частичного ограничения панмик-сии способствуют закреплению этих наиболее удачных комбинаций. Если организм вполне приспособлен к известным условиям существования и эти условия заметно не меняются на данном этапе истории, то его строение может длительное время оставаться почти неизменным. Однако эта неизменность лишь кажущаяся. Под покровом определенного приспособленного фенотипа идет непрерывная перестройка наследственной сущности организма на основе указанных процессов мутирования, размножения и комбинирования мутаций под контролем естественного отбора. Эти изменения могут иметь скрытые формы накопления более или менее значительного резерва изменчивости внутри популяции. Они могут приобрести и более явные формы при изменении зависимостей организма от факторов внешней среды (стабилизация и лабилизация формообразования). Но могут привести и к более или менее значительному изменению фенотипа организма, к изменению его индивидуального развития, его функций, а следовательно — и его положения во внешней среде.
Современная генетика оперирует не с «признаками», а только с «нормой реакций» организма и ее наследственными изменениями. Введение понятия «нормы реакций», охватывающего как строение, так и функции организма во всем его онтогенезе, имело и имеет огромное значение в развитии биологии. Именно в таком широком смысле понятие «нормы» приобретает исчерпывающую определенность, совершенно необходимую для точного анализа наследственных изменений.
Однако при рассмотрении эволюционных проблем приходится понятие «нормы реакций» дифференцировать. Для нас имеют особое значение, с одной стороны, та в известной мере стойкая «типичная» организация (включая ее функции), с которой оперирует систематика и которая является результатом более или менее автономных процессов развития, определяемых внутренними, наследственными факторами, и, с другой стороны, более или менее широкая индивидуальная изменяемость или лабильность той же организации, определяемая известной зависимостью процессов онтогенеза от изменений в факторах внешней среды. В первом случае мы имеем дело с реакциями развивающегося организма на внутренние факторы развития (внешние факторы играют роль условий нормального развития). Во втором случае — с реакциями организма на внешние факторы развития (имеющие до известной степени детерминирующее значение). В первом случае речь всегда идет о более или менее сложных зависимостях внутри развивающегося организма, т. е. о корреляциях в широком смысле (включая генетические). Особое значение имеют в эволюции корреляции регуляторного характера. Во втором случае вопрос касается способности организма отвечать на изменения факторов внешней среды изменением своей организации и своих функций. Можно говорить о зависимых процессах или о реакциях в узком смысле, и о реакционных механизмах организма. Наибольшее значение имеют в эволюции реакции приспособительного характера. Поэтому мы должны обратить внимание, с одной стороны, на источники возникновения новых корреляций и на эволюцию корреляционных механизмов регуляторного значения и, с другой стороны, на источники возникновения новых реакционных механизмов и на эволюцию их адаптивности.
В основе процесса эволюции лежат наследственные изменения отдельных особей — мутации.
В конкретных условиях развития устранение одних выражений мутаций и усиление других означает изменение всей системы реакций организма. Изменяются выражения, зависящие от факторов внешней среды, и изменяются также те выражения, которые определяются в основном внутренними факторами развития. Множественность выражения мутаций (плейотропия) означает наличие известных зависимостей в изменениях различных признаков и реакций. Следовательно, изменение различных выражений всегда связано с изменениями в этих зависимостях. Этим самым создается база для возможных перестроек систем корреляций в процессе эволюции организма.
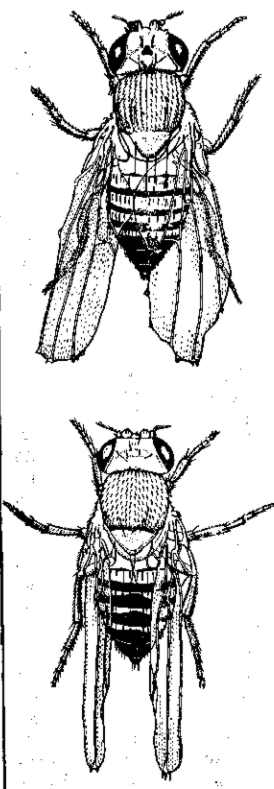
Рис. 19. Мухи Drosophila melanogaster из чистой линии beaded. Вверху — среднее выражение признака, внизу — наибольшее выражение. По Г. Мёллеру, 1918
1. ИЗМЕНЕНИЕ ВЫРАЖЕНИЯ МУТАЦИЙ В ПРОЦЕССЕ ЭВОЛЮЦИИ
Не только мута|бильность (мутационное «давление»), но и отборное значение, коэффициент селекции) признаков не остаются постоянными. И это объясняется не только постоянными изменениями факторов среды, но и изменениями самого организма. Даже отдельная мутация не имеет постоянного выражения (рис. 19). Поэтому и концентрация мутаций не устанавливается на каком-либо уровне равновесия. Этот уровень непрерывно меняется, по меньшей мере — для всех небезразличных признаков.
Так как отдельные мутации вносят обыкновенно неблагоприятные изменения организации (с ее функциями), то нелегко было бы понять, каким образом возможна прогрессивная эволюция, если бы мы не владели громадным арсеналом фактов, показывающих, что неблагоприятные выражения мутаций в процессе эволюции погашаются и преобразуются. Это достигается в результате комбинирования различных мутаций при скрещиваниях и естественном отборе наиболее жизненных сочетаний.
Генетиками уже давно было установлено, что фенотипическое выражение мутаций может резко изменяться при их комбинировании. Классическим примером такого комбинирования являются различные формы гребня у петухов (рис. 20). В данном случае, как и во многих других, наблюдается именно коренное преобразование выражения единичных мутаций. Комбинация дает качественно новое формообразование. В других случаях мутация получает известное выражение вообще лишь в комбинации с определенной другой мутацией — в отдельности мутация не проявляется. Такие факторы генетики называют дополнительными («комплементарными»). Так, например, основной «ген» окраски у домашней мыши определяет развитие серой или черной окраски лишь в присутствии дополнительных факторов. Известны также дополнительные факторы, определяющие развитие окраски цветов душистого горошка. Наконец, в третьих случаях, наоборот, мутация полностью теряет свое выражение при комбинировании с определенной другой мутацией, которая оказывает «эпистатическое» влияние на первую. Классическим примером такого эпистаза может служить доминантная белая окраска итальянских кур (леггорнов), которая подавляет развитие иных окрасок.
Эти примеры относятся к крайне резкому проявлению эффекта комбинирования двух простых мутаций. Выражение одной мутации проявляется лишь при наличии другой (комплементарной) или, наоборот, полностью подавляется (эпистатическим фактором). Все ото указывает на наличие известного взаимодействия, конечно — не непосредственно между «генами», как это иногда упрощенно описывают, а между теми биохимическими и морфогенетическими процессами, которые оказались измененными в результате данных мутаций.

Рис. 20. Гребни домашних кур. Доминирование уклоняющихся форм гребня — гороховидного (А) и розовид-ного (В)—над простым (С). Комбинирование гороховидного и розовидного гребней дает качественно новую ореховидную (D) форму гребня. По Р. Пеннет и П. Бейли, 1921
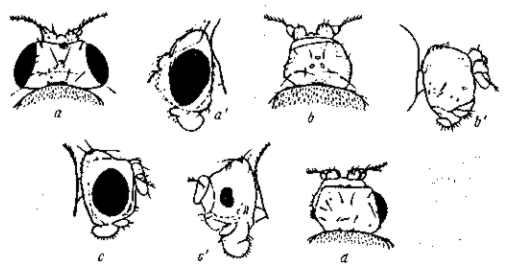
Рис. 21. Изменчивость выражения мутации безглазия (eyeless) у Drosophila melanogaster. а, а' — нормальные глаза; b, b' — совершенно безглазая форма. В чистой культуре в течение ряда поколений выражение этой мутации постепенно теряется (с, с') вплоть до восстановления почти нормальных размеров глаза) и нормальной жизнеспособности мутантных мух. По Т. Моргану, 1916
Мы сослались на известные всем генетикам типичные примеры такого взаимодействия. В некоторых случаях модифицирующее влияние одних мутаций на другие выражается исключительно ярко. В менее резкой форме подобное взаимодействие не составляет исключения, нуждающегося в определенных терминах, а является обычным правилом. Меняется выражение и в компаундах разных аллеломорфных мутаций. Иногда, правда, бывает, что каждая мутация сохраняет свои особенности и в компаунде (например, aristopedia X spineless). Чаще, однако, эти выражения меняются. Интересно, что нередко выражения обоих аллеломорфов взаимно погашаются. Так, vortex (вихреобразные щетинки) и oblique (косо срезанные крылья) дают в компаунде почти нормальный фенотип. То же самое касается split bristles X Notch, а также achaete X scute-1, теряющих в компаунде свои выражения (Мёллер).
«Доминантная» мутация четковидных крыльев (Beaded) обладает в гетерозиготном состоянии крайне изменчивым выражением, меняющимся как при комбинировании, так и при изменении внешних факторов (рис, 19). Фактор black (черное тело) даже в гетерозиготном состоянии подавляет до известной степени выражение Beaded, а мутация vestigial (рудиментарные крылья) в гетерозиготном же состоянии усиливает его эффект. Мутация sooty оказывает также усиливающее влияние на выражение мутации четко-видных крыльев (Мёллер). При выращивании личинок этой мутации в подсохшей среде четковидность оказывается менее выраженной и часто вовсе не проявляется, так что значительная часть мух оказывается фенотипически нормальной. Так же влияют и различные другие внешние факторы (Декстер, 1914). Большая зависимость от генотипа установлена и для выражения мутации Beaded в гетерозиготном состоянии (Шифрин, 1941).
Рецессивные мутации рудиментарных крыльев (vestigial), а также безглазия (eyeless) значительно изменяются в зависимости от температуры (рис. 21, 22) и от качества корма. Мутация безглазия также весьма чувствительна и по отношению к модифицирующим влияниям со стороны других мутаций. Однако иногда изменчиво даже выражение «доминантной» мутации в гомозиготном состоянии. Так, мутация уродливого брюшка (Abnormal abdomen) дрозофилы в своем выражении весьма зависима от влажности корма, а мутация полосковидных глаз (Ваг) весьма чувствительна к изменениям температуры (рис. 6).
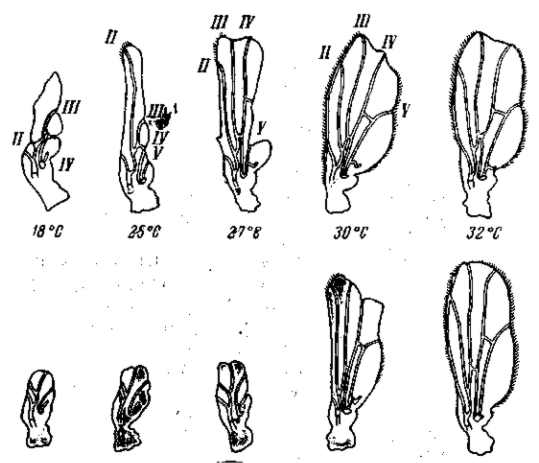
Рис. 22. Зависимость формы и величины крыльев мутации vestigial (Drosophila теlanogaster) от температуры Верхний ряд — самцы, нижний — самки. При 32° в культуре этой мутации 18% самцов и 15 % самок имеют совершенно нормальные крылья (проявление становится неполным). По М. Г. и M. Л. Харнли, 1935
Как мы раньше уже отмечали, каждая мутация выражает либо изменение времени, скорости образования и концентрации морфо-генных веществ, либо изменение реактивности тканей (сдвиг пороговых уровней нормальных реакций). В обоих случаях это означает взаимное смещение формативного раздражителя (активатора) и реагирующей системы (реактора). В огромном большинстве случаев смещение приводит также к уменьшению «морфогенного запаса» и приближению измененной интенсивности активатора (индуктора), к одному из порогов нормальной реактивности реактора. Такое приближение естественно ведет к уменьшению устойчивости формообразовательного процесса. Близость порогового уровня означает незащищенность морфогенетической реакции достаточным «запасом». Самое незначительное дальнейшее уклонение в том же направлении,— либо под влиянием небольших наследственных изменений (малых мутаций), либо под влиянием изменения внешних факторов,— может привести к превышению порога нормальной реактивности и к более или менее значительному нарушению формообразования. Поэтому те мутации, которые стоят как раз на грани подобных нарушений, оказываются особенно неустойчивыми в своем проявлении (т. е. обладают неполным проявлением) или в своем выражении.
Полностью рецессивные мутации в гетерозиготном состоянии не проявляются, т. е. в однократной «дозе» при наличии нормального аллеломорфа вызывают сдвиг, не достигающий порогового уровня нормальной реактивности тканей. В гомозиготе они проявляются, и это указывает на то, что двукратная «доза» изменения «гена» выводит взаимодействующую морфогенную систему за пределы порога нормальной реактивности. Если она оказывается как раз на грани этого порога, то такая мутация будет в своем проявлении и выражении весьма неустойчивой. Она окажется в широких пределах зависимой как от наличного генотипа, так и от внешних влияний. Возможна мутация, которая не проявляется и в гомозиготе, но все же вызывает заметный сдвиг нормальной реактивности, который при суммировании с другой мутацией приведет к видимому изменению фенотипа или к изменению его физиологического состояния. Это — «малая» мутация, выступающая лишь в роли «модификатора».
С другой стороны, возможны более эффективные мутации, которые не только в двукратной дозе, но и в гетерозиготе, при наличии нормального аллеломорфа, достигают или даже превышают допустимые пределы сдвига компонентов взаимодействующей системы. Если сдвиг достиг порогового уровня, то получится картина доминирования с неполным проявлением (в зависимости от генотипа и внешних факторов). Если этот пороговый уровень полностью превзойден, то возможно устойчивое проявление данной мутации при изменчивости ее выражения.
Ни об одной мутации не известно, чтобы она достигла того уровня устойчивости, каким обладает нормальный организм. Это означает, что если порог «нормальной» реактивности и превзойден полностью (как в доминантных мутациях), то все же не настолько, чтобы создать «запас», «защищающий» формообразование мутации. Такие «запасы» создаются исключительно в процессе эволюции (т. е. при направляющем влиянии отбора), а не в порядке единичных мутаций. Они «защищают» развитие уже установившейся «нормы» («фактор безопасности» Холдена).
Мутации никогда, по-видимому, не достигают уровня полной доминантности (пока они остаются в роли мутаций). То, что у мутаций дрозофилы называется доминированием, никогда не достигает высоты доминирования нормального фенотипа. Доминирование нормы выражается обычно в почти полной или полной неотличимости гетерозиготы от гомозиготы (по фенотипу). Между тем при доминировании мутации гетерозигота всегда легко отличима от гомозиготной мутации. Последняя обладает гораздо более ярким выражением характерных для нее признаков. Гетерозигота доминантной мутации занимает всегда промежуточное положение между нормой и гомозиготной мутацией. Таковы все известные доминантные мутации дрозофилы. Одним словом, доминантные мутации на самом деле всегда бывают полудоминантными.
Полудоминанты отличаются неустойчивостью именно потому, что они не достигли уровня настоящих доминантов, т. е. той активности, которой отличаются «нормальные» аллеломорфы. Вернее — измененные в мутациях формообразовательные процессы никогда не обладают той «защищенностью», которая характерна для процессов индивидуального развития нормального «дикого» типа. Это связано с кратковременностью их исторического существования и отсутствием положительного селекционного значения данной мутации.
Мы видим, что проявление и выражение мутаций, как в гетеро-зиготе, так и в гомозиготе, суть величины переменные. Они могут изменяться как в сторону усиления, так и в сторону ослабления. В этом смысле имеют значение и внешние факторы среды, в которой развивается организм (мутация), и внутренние факторы, т. е. генотип данной особи.
В процессе эволюции особое значение имеет комбинирование. При скрещиваниях создаются все новые комбинации, а вместе с тем меняются выражения мутаций, меняются фенотипы и их реакции (рис. 15, 16). Так как большинство мутаций так или иначе неблагоприятно, то наибольшее значение имеет частичное или полное погашение их выражения.
А. ОБЕЗВРЕЖИВАНИЕ НЕБЛАГОПРИЯТНЫХ МУТАЦИЙ (ЭВОЛЮЦИЯ РЕЦЕССИВНОСТИ)
Неоднократно делались попытки повлиять путем искусственного отбора на мутации с изменчивым выражением. Эти попытки давали положительные результаты. Путем искусственного отбора удавалось значительно поднять выражение мутации до известного максимума и удавалось снизить его, иногда — до уровня дикой нормы, т. е. до полной утраты проявления. Эти изменения оказывались наследственными и зависели, следовательно, от отбора небольших наследственных изменений (малых мутаций), меняющих выражение культивировавшихся мутаций. Такие малые мутации были названы «модификаторами». В этом «модифицировании», конечно, нет ничего специфического,— мы ведь видели, что взаимодействие «генов» (точнее — измененных биохимических и морфо-генетических процессов) вообще связано с изменением выражения отдельных мутаций. В термине «модификатор» подчеркивается, однако, модифицирующее влияние только потому, что во многих малых мутациях нам не удается пока уловить их влияния на нормальный организм (так как «дикая» норма отличается гораздо большей устойчивостью, т. е. «защищенностью» онтогенеза). Поэтому их наличие пока устанавливается только по влиянию на выражение мутаций и то главным образом лишь по комплексному влиянию целого ряда малых мутаций.
Анализ фенотипов любой природной популяции показывает, обычно, поразительное единообразие. В частности, в популяциях различных видов дрозофилы, в том числе и классической Drosoрhila melanogaster, решительно господствует нормальный «дикий» тип. И все же генетический анализ показывает огромную их насыщенность разнообразнейшими мутациями, в том числе и полудоминантными. Все эти мутации находятся, однако, в скрытом виде, большей частью в качестве рецесссивов (в гетерозиготном состоянии; С. Четвериков, Н. Дубинин и др.). Однако встречаются не так уж редко и полудоминанты — выражение их частью подавлено эпистатическими влияниями генотипа или же они обладают низким процентом проявления (Р. Берг, С. Гершензон). Встречаются и довольно ясно выраженные мутации (по Р. Берг, особенно много полудоминантных аутосомных мутаций) и притом не безвредные. Имеется в природных популяциях и немало летальных (Н. Дубинин, Ф. Добжанский), а также стерильных (Р. Берг, 938) мутаций. Однако в особенности велико насыщение малыми мутациями, выступающими в роли модификаторов (Н. Дубинин и др.), и физиологическими мутациями, отражающимися лишь на относительной жизнеспособности (Г. Муретов, 1941). Это показывает, что в природных условиях происходят те же процессы насыщения популяций мутациями и их обезвреживания путем отбора наиболее благоприятных комбинаций, т. е. прежде всего отбора наиболее «жизнеспособных» особей, какие установлены для лабораторных культур.
И в природных условиях неблагоприятные мутации теряют свое выражение. В основном это касается гетерозиготы, так как гомозиготные мутации могут встретиться в природе лишь в виде исключения. Результатом утраты своего выражения в гетерозиготе является рецессивность мутации.
То, что нами наблюдается в лаборатории и в природных условиях, является лишь ничтожным отрезком истории организмов. Те мутации, которые мы культивируем как вновь возникшие наследственные изменения и на которых мы наблюдали изменение их выражения в течение исключительно короткого времени, возникали многократно и ранее в истории данного вида организмов (например, дрозофилы). Организм уже до известной степени приспособился к ним, т. е. выработал различные компенсации, подавляющие их неблагоприятные выражения. В частности, под этим углом зрения можно оценивать обычные явления доминирования нормы и рецессивности мутаций.
В действительности, нормальный фенотип более или менее полно доминирует над всеми мутациями. Мы уже отмечали, что даже немногие известные «доминантные» мутации дрозофилы в действительности являются полудоминантными. Их выражение в гетерозиготе всегда ослаблено по сравнению с гомозиготой. Частично это может объясняться тем, что большинство мутаций связано с ослаблением интенсивности нормальных процессов развития (замедлением продукции и снижением концентрации морфогенных раздражителей или снижением реактивности тканей, т. е. поднятием нижнего порога и снижением верхнего порога чувствительности) и ведет к известному недоразвитию или упрощению морфогенеза. Генетики говорят об инактивировании «генов» и «гшгоморфных» или «аморфных» генах (Мёллер, 1932).
Однако уже факт существования обратных мутаций показывает возможность противоположных изменений в сторону интенсификации морфогенетических процессов («гиперморфные» гены Мёллера — ускорение образования и повышение концентрации формообразовательных веществ, а также повышение чувствительности тканей, т. е. раздвигание обоих пороговых уровней их реактивности). Возможны и такие изменения, которые противодействуют течению нормальных реакций или вносят в них нечто совершенно новое (антиморфы и неоморфы Мёллера). В этих случаях наблюдается доминирование мутации. Однако, как уже сказано, это доминирование в действительности никогда не бывает полным. Можно предполагать, что выражение этих мутаций также уже частично ослаблено. Вместе с тем нельзя не отметить, что таких мутаций известно очень немного. Огромное большинство мутаций дрозофилы — рецессивно. Очевидно, это объясняется так же, как и в прошлой истории вида: при повторном возникновении этих, не слишком вредных, мутаций шел естественный отбор комбинаций, нейтрализующих их выражение. Такие комбинации (генотипы) отличались большей жизнеспособностью и распространялись в популяции в качестве все более устойчивой «нормы».
Одновременно с развитием генетического механизма, обезвреживающего наиболее частые мутации, происходило упрочение процессов нормального формообразования даже гетерозиготного организма, т. е. усиление доминирования нормы, и вместе с тем ослабление выражения неблагоприятных мутаций (в той же гетерозиготе) вплоть до их полного погашения, т. е. приближение к полной рецессивности мутаций.
Теория эволюции доминантности Фишера построена на значении отбора малых мутаций с модифицирующим влиянием. Предполагается, что каждая действительно новая мутация обладает всегда некоторым выражением и в гетерозиготном состоянии, т. е. все вновь возникающие мутации полудоминантны. Если такая мутация непосредственно не элиминируется, т. е. по своему выражению не слишком неблагоприятна (при данных условиях существования), и оставляет какое-то потомство, то она комбинируется в этом потомстве с несколько различными генотипами. Выражение ее оказывается изменчивым, и в процессе естественного отбора переживают всегда наиболее приспособленные, наиболее жизнеспособные комбинации. Если такой процесс многократно возобновляется и поддерживается мутированием и распространением мутации, то постоянный отбор в пользу нормального фенотипа приводит к накоплению в данной популяции «модификаторов», нейтрализующих вредные выражения распространенных мутаций. Тем самым эти мутации становятся рецессивными в данной популяции, насыщенной соответствующими модификаторами, а норма — по отношению к ним — доминантной.
Если среди новых полудоминантных мутаций встречается относительно (в данных условиях) благоприятное изменение, то его выражение будет через тот же отбор «модификаторов» (т. е. попросту через отбор более приспособленных особей) постепенно усиливаться, вплоть до максимально возможного выражения в гетерозиготе. Это означает приобретение полной доминантности.
Благоприятная доминанта быстро распространится при участии положительного отбора, перейдет в гомозиготное состояние и войдет в состав новой нормы, вытесняя прежнюю норму; прежний нормальный аллеломорф потеряет свою доминантность и в качестве рецессива перейдет в скрытое состояние.
Таким образом, большинство не слишком вредных мутаций становится рецессивным. Однако такие мутации, которые заметно ослабляют жизнеспособность и приспособленность организма к данным условиям внешней среды, не имеют шансов переживать и оставлять потомство. Ясно, что такие постоянно элиминируемые мутации сохраняют всегда свои исходные свойства «новых» мутаций, т. е. остаются полудоминантными, так как никакой отбор среди нежизнеспособных особей невозможен. В результате целый ряд мутаций дрозофилы оказывается полудоминантным. В особенности это касается ряда более значительных физиологических мутаций с летальным или стерильным эффектом.
Другие авторы, присоединяясь к основным положениям Фишера об эволюции доминантности, представляют себе этот процесс несколько иначе. Райт, Холден и Мёллер считают, что доминантность выражает активность факторов нормального развития. Обычно при этом говорят об активности нормальных «генов» и об инактивации мутантных (в большем или меньшем согласии с представлениями Гольдшмидта о количественном характере изменений генов и об их энзиматозной природе). Нам незачем связывать вполне реальные данные об изменении активности или концентрации формативных веществ и об изменении времени и скоростей морфогене-тических реакций с гипотетическими представлениями о природе гена, о характере его изменений и о способе его участия в индивидуальном развитии организма. Я пытался показать (И. Шмальгаузен, 1938а, 1942), что почти все мутации, вызывая сдвиги в мор-фогенетических системах (между «активатором» и «реактором») организма, вносят известное нарушение развития, которое при превышении порогового уровня известной реакции вызывает выпадение этой реакции. Такие выпадения некоторых реакций должны выражаться в большей или меньшей степени недоразвития. Обычно мутация как раз и связана с известным недоразвитием, т. е. с остановкой развития или заменой сложного онтогенетического процесса более простым. Естественно, что такие мутации имеют характер «гипоморфньгх» мутаций и в комбинации с нормальным аллеломорфом оказываются до известной степени рецессивными. Однако большая устойчивость нормы и более или менее полная рецессивность, очевидно, не объясняются так просто.
По мнению Мёллера, понижение (или повышение) степени выражения известной мутации зависит не от подбора генов — модификаторов этой мутации, а от отбора на большую (или меньшую) активность того же основного гена, который определяет развитие нормы. При наличии ряда аллеломорфных изменений, градуально друг от друга отличающихся, такой отбор вполне возможен. Следовательно, вредные мутации будут уничтожаться через отбор гетерозиготных особей с наименьшим их выражением, т. е. наибольшей активностью «нормального» аллеломорфа. Наоборот, благоприятные мутации будут повышать свою активность, т. е. увеличивать степень доминирования. Повышение активности до известного уровня приведет к тому, что уже один ген (1А) даст полную реакцию в виде реализации характерных признаков в их максимальном (оптимальном) выражении, так что его удвоение (2А в гомозиготе) не даст уже никакого дополнительного эффекта. В таком случае будет достигнуто полное доминирование. Те мутации, которые имеют положительное значение для жизни вида, будут всегда подбираться по этому пути повышения активности вплоть до развития полной доминантности. Полезная мутация, став доминантной, распространится с большой скоростью по всей популяции и войдет в состав ее нормы.
Мутации, до известной степени вредные, будут переходить в менее активное состояние и в обезвреженном виде могут сохраняться в популяции в виде скрытого резерва ее изменчивости (в гетерозиготах).
По представлениям Райта, степень доминирования есть также одно из выражений активности гена, и рецессивность означает его переход в менее активное состояние. Холден не отрицает значения модификаторов, действующих на мутацию. Холден добавляет к этому, однако, еще и следующие соображения. Если нормальный ген вызывает уже в гетерозиготе полный эффект, то всякое увеличение его активности останется незамеченным. Мутация с повышенной активностью («гиперморфы» Мёллера), которые могли бы быть полностью доминантными, не получают видимого выражения. Только минус-мутация может получить свое выражение, если активность гена в гетерозиготе падает ниже нормы, т. е. если она рецессивна. Такие мутации обычно менее жизнеспособны и частью элиминируются, частью видоизменяются в процессе эволюции. С другой стороны, плюс-мутации, связанные с переходом «гена» в более активное состояние, хотя и не дают видимого эффекта, но ведут к увеличению устойчивости нормы. Они свободно накапливаются в процессе отбора. Поэтому отбор всегда будет действовать в сторону повышения активности (и доминирования) всех жизненно важных генов.
Исследования результатов межвидовой гибридизации некоторых видов хлопчатника показали, что доминантность имеющихся у них параллельных мутаций зависит от различных (у разных видов) комплексов модификаторов. Они приобретены в истории данных видов независимо друг от друга и имеют поэтому разный состав. При межвидовом скрещивании доминирование оказывается лишь слабо выраженным вследствие очевидной дезинтеграции обеих систем модификаторов, определяющих это доминирование (Харланд, 1936, 1937). В особенности ясно это проявляется при внесении мутации в чуждый ей вид путем повторного возвратного скрещивания с этим видом. Это является прекрасным доказательством правильности теории эволюции доминантности в том ее виде, как она была разработана Фишером (1931). Доказательствами эволюции рецессивности являются и следующие данные того же Харланда. Фактор crinkled — dwarf оказывается простым рецессивом в одном сорте хлопка, однако в скрещивании с другими сортами хлопка, т. е. в других комбинациях генов (включая «модификаторы»), он обнаруживает во втором поколении, полученном путем самоопыления гибридов, все степени доминирования. По Фишеру, это объясняется расщеплением в отношении многочисленных модификаторов. Однако и в этих случаях допустимо не только предположение о дезинтеграции системы модификаторов, обусловливающих рецессивность данной мутации. Можно допустить и другие толкования, в особенности потому, что эта же мутация оказывается вполне рецессивной и в хлопчатнике «упланд» (Gossypium hirsutum), в котором она никогда не встречается и который, по Фишеру, не может обладать системой модификаторов, обусловливающих ее рецессивность.
В данном случае эти детали не имеют большого значения, и нам представляется более правильным считать, что как в процессе эволюции вообще одинаковые результаты достигаются самыми различными средствами, так и в данном случае весьма вероятно, что эволюция рецессивности (и доминантности) идет разными путями. Повышение устойчивости нормы как путем отбора модификаторов, так и через отбор более активных аллеломорфов, при которых достигается наибольшая надежность формообразования не только в гетерозиготе, но и в гомозиготе, представляет одну сторону общего процесса стабилизации нормы. Уменьшение выражения всех вредных мутаций в результате отбора менее активных аллеломорфов, а также отбора модификаторов и вообще различных наследственных сочетаний представляет другую сторону того же процесса стабилизации нормы, который осуществляется всегда и повсюду через стабилизирующую форму естественного отбора на основе селекционного преимущества нормы перед всеми от нее уклонениями.
Б. ИЗМЕНЕНИЯ ЧАСТИЧНО ВРЕДНЫХ МУТАЦИЙ (ВОЗНИКНОВЕНИЕ КОРРЕЛЯЦИЙ)
Так как мутации имеют обычно множественное выражение, а благоприятные выражения встречаются исключительно редко, то большое значение в эволюции имеет возможность нейтрализации одних лишь вредных выражений, а также возможность качественного преобразования других выражений и усиления положительного эффекта тех же мутаций. Такие сложные преобразования предполагают существование весьма дифференцированного действия модификаторов и приводят к установлению сложных систем связей (генетических корреляций), входящих тогда как необходимый элемент наследственной структуры организма.
Возможность дифференцированного изменения различных выражений одних и тех же частично вредных мутаций доказывается целым рядом фактов. Напомним прежде всего, что в основе мутаций лежат, очевидно, некоторые биохимические изменения, оказывающие влияние на процессы внутриклеточного обмена веществ. Только через эти изменения в обмене веществ развивающегося организма они вызывают нарушения в ходе морфогенетических процессов и приводят к видимому изменению нормального строения организма. Поэтому нужно думать, что любая морфологическая мутация имеет обязательно и физиологическое выражение. Даже малые мутации, не проявляющиеся в заметных изменениях структуры (хотя бы и в силу наличия совершенной регуляции), могут все же иметь немалое физиологическое значение. На самом деле, мы знаем, что все «вредные» мутации дрофозилы обладают ослабленной жизнеспособностью, пониженной плодовитостью, измененным темпом роста или развития, измененной реактивностью или другими физиологическими выражениями. С другой стороны, действительно известны как малые, так и большие физиологические мутации, не обладающие видимым морфологическим выражением.
Целый ряд видимых мутаций дрозофилы связан с резким снижением жизнеспособности — они «летальны» или полулегальны. Не может быть случайностью, что такие мутации оказываются в своем морфологическом выражении обычно полудоминантами, а в физиологическом выражении — рецессивами. Таковы «доминантные» мутации Notch, Dichaete, Star, Beaded, Truncate. Все они в гетерозиготе вполне жизнеспособны, а в гомозиготе оказываются полностью летальными. То же самое найдено и у амбарной огневки (Ephestia kuhniella). Мутация, сдвигающая поперечные полосы рисунка крыльев (Sy), вполне выражена, т. е. полудоминантна в гетерозиготе, и вполне жизнеспособна. Однако в гомозиготе она детальна. Известная мутация желтой окраски у домашней мыши доминантна, но обладает рецессивным летальным действием, обнаруживающимся в гомозиготе. Коротконогость кур (криперов) доминантна, но в гомозиготе она детальна. То же самое касается известных пород домашнего скота Niatta и Dexter. Последние обладают свойствами, ценимыми человеком, и потому здесь можно говорить лишь о влиянии искусственного отбора. Что же касается дрозофилы и огневки, то различная активность (доминантность) разных выражений одних и тех же мутаций является, очевидно, результатом естественного отбора.
По-видимому, морфологические выражения этих мутаций были не столь вредны, как ослабление общей жизнеспособности в гетерозиготном состоянии. Кроме того, постоянное наличие большого запаса малых «физиологических» мутаций способствовало большой эффективности отбора на «жизнеспособность» гетерози-готы, т. е. на устранение проявления признаков «летальности». Поэтому физиологическое выражение этих мутаций снижено до полной рецессивности, в то время как менее вредное морфологическое выражение осталось еще полудоминантным. Во всяком случае, это показывает возможность изменения соотношений разных выражений одной и той же мутации в гетерозиготе и в гомозиготе.
Несомненно, возможны были бы и диаметрально противоположные соотношения — погашение морфологического выражения при сохранении физиологического. Это доказывают результаты некоторых опытов комбинирования мутаций у той же дрозофилы.
Мутация lozenge обладает грубыми фасетками неправильной формы, а мутация spectacled характеризуется блестящей поверхностью глаза без фасеток. Это — аллеломорфы, которые, каждая в отдельности, нарушают строение глаза, но, кроме того, связаны с бесплодием самок. В компаунде обеих мутаций глаза получают нормальное строение, однако самки остаются бесплодными. Аллело-морфные мутации split bristles и facet также нарушают строение омматидий. В компаунде восстанавливается нормальное строение глаз, но развиваются добавочные щетинки (как в первой мутации). Черночешуйчатая мутация амбарной огневки (Ephestia kiihniella) рецессивна, но снижение жизнеспособности (на 22%) у нее доминантно (случай, обратный обычному; возможно, что это особая форма защиты организма от вредной мутации; тогда она могла развиться лишь в процессе межгрупповой конкуренции разных популяций) . Эти факты показывают возможность изменения отдельных выражений мутаций в различных направлениях через процесс их комбинирования.
Добжанский (1927) указывает, что аллеломорфы белых глаз (white), а также мутации окраски тела sooty и ebony у дрозофилы рецессивны, поскольку более выраженная неблагоприятная окраска у гетерозиготы, очевидно, элиминировалась и в процессе отбора произошло восстановление нормальной, дикой окраски. Однако плейотропное выражение тех же мутаций — измененная форма сперматеки, очевидно (как нейтральный признак внутренней организации) , не была предметом отбора. Она оказывается у них полудоминантной и дает в гетерозиготе промежуточные формы.
Хохлатость кур связана с гидроцефалией и недоразвитием крыши черепа. Однако эти последние выражения очень изменчивы и могут быть подавлены, как это видно из факта существования пород хохлатых кур, имеющих нормальные черепа.
Мохноногость кур связана с брахидактилией четвертого пальца. Однако и это выражение очень изменчиво, что указывает на существование многих модификаторов. В самом деле, брахидактилия может быть полностью подавлена. У некоторых пород кур (фаве-ролей) мохноногость не сопровождается брахидактилией.
В случае положительного отбора по одному выражению и элиминации особей с более значительными иными выражениями возможна, таким образом, стабилизация положительного и устранение отрицательных выражений. Это происходит через отбор мутаций с модифицирующими влияниями. Возможно установление новой нормы, обладающей известными преимуществами перед прежней. В этом случае новый благоприятный признак связан с целой системой модификаторов, совместно участвующих в его развитии. Они становятся необходимыми элементами аппарата индивидуального развития. Они действуют притом на одни биохимические реакции и на одни части морфогенетической системы как активаторы, а на другие части как депрессоры или трансформаторы реакций. Это означает установление сложных связей между различными процессами. Если уже самый факт наличия плейотропного (множественного) выражения мутации показывает наличие известных коррелятивных зависимостей между физиологическими и морфогенетическими процессами, то преобразование различных выражений этой мутации путем комбинирования со многими модификаторами означает укрепление и преобразование данной системы корреляций, а также создание новой системы геномных связей. Развитие отдельных признаков оказывается зависящим не только от основного мутант-ного «гена», но и от многих других наследственных факторов. Это выражается в наличии сложных связей между биохимическими процессами, определяющими наступление известных формообразовательных реакций, а также в существовании взаимозависимостей между этими последними.
В процессах преобразования выражения мутаций происходит поэтому стабилизация процессов формообразования через развитие системы геномных и морфогенетических корреляций, связывающих организм в более устойчивое целое (И. Шмальгаузен, 1938а, 1942). При этом развитие нормы становится более защищенным не только от небольших мутаций, но и от внешних влияний. Руководящим фактором в этих процессах является стабилизирующая форма естественного отбора (т. е. постоянная элиминация неблагоприятных уклонений).
В. ИЗМЕНЕНИЯ ПРОЯВЛЕНИЯ МУТАЦИЙ (ВОЗНИКНОВЕНИЕ ГЕТЕРОМОРФИЗМА И ПОЛИМОРФИЗМА)
Если мутация с изменчивым выражением колеблется между ясно видимым проявлением и полным его отсутствием, то говорят о неполном ее проявлении. Неполным может быть проявление полудоминантной мутации в гетерозиготном состоянии, а также рецессивной, а иногда даже полудоминантной мутации в гомозиготе. Неполное проявление указывает на особую лабильность данной мутации, на резко выраженную зависимость формообразования от наследственных свойств каждой особи и притом именно от ничтожных различий в генотипе, обусловленных присутствием тех или иных малых мутаций (модификаторов). Вместе с тем лабильность мутации выражается и в не менее резко выраженной зависимости формообразования от изменений в факторах внешней среды (температуры, влажности, корма и т. п.). Это служит основой развития генотипического и фенотипического разнообразия форм внутри популяции.
Так как изменчивость выражения и проявления указывает на значительную зависимость развития данных признаков (мутантов) от небольших генотипических различий, а также от изменений в факторах внешней среды, то этим самым дана возможность быстрого изменения этого выражения в результате естественного отбора. Если мутация неблагоприятна, то ее выражение должно бы исчезнуть в результате стабилизирующего отбора в пользу нормы. Если же она благоприятна, то она должна бы быть усилена до полной доминантности и войти в состав нормы в результате ведущей формы естественного отбора.
Если это не происходит с целым рядом особенно изменчивых мутаций, то для этого должны быть свои основания. Очевидно, в природной обстановке имеются условия, поддерживающие эту изменчивость. В пользу этого говорит и распространенность полудоминантных мутаций с неполным проявлением в природных популяциях (С. Гершензон, 1941а, б). При этом доминантность таких мутаций выражается сильнее при скрещиваниях внутри своей популяции, чем при скрещивании с другими популяциями (Р. Берг). Это означает отсутствие отбора на снижение их выражения в гетерозиготе, а быть может, даже свидетельствует о наличии положительного отбора. Можно думать, что существует система модификаторов, усиливающих выражение этих мутаций, и что эта система при скрещиваниях с другими популяциями частью разрушается.
Быть может, в таких мутациях их морфологическое выражение не столь неблагоприятно, как это нам кажется, и можно предполагать наличие прочной (т. е. неустранимой) корреляции с какими-либо положительными изменениями физиологического характера. В таком случае мы имеем лишь частично вредные (частично полезные) мутации.
Распространенность таких мутаций в больших популяциях заставляет также думать, что во многих случаях она относится к группе лишь условно вредных мутаций. Широкое распространение популяции, сталкивающейся с разнообразными микрогеографическими условиями, позволяет предполагать, что некоторые мутации могут оказаться в относительно благоприятном положении в определенных частных условиях существования (С. Гершензон) (или в известных сезонах, или при случайных изменениях в факторах среды). В этом случае концентрация мутации будет поддерживаться на известном уровне соответственно частоте встречаемости данных частных условий существования. Возможно, что как гетерозигота, так и обе гомозиготы в процессе эволюции разовьют свои условно наиболее благоприятные выражения. Возможно, наконец, и установление более сложных, дигибридных комбинаций со многими, вполне в известных условиях приспособленными формами.
Это приведет к установлению постоянного гетероморфизма данного вида. Частными случаями такого гетероморфизма являются различные окраски богомолов и различные формы мимикрии самок у Papilio polytes и P. dardanus. В других случаях в популяциях встречаются и безусловно вредные, но все же «доминантные» мутации. Как уже упоминалось, в таких случаях можно думать о доминантности как об особой форме «защиты» популяции от распространения вредной мутации. Такая «защита» могла (по Р. Л. Берг) развиться лишь в результате межгрупповой конкуренции, в которой побеждали более «здоровые» популяции. Однако мы видели, что мутации, по Фишеру, возникают как полудоминантные. Элиминируемые мутации не могут менять в процессе эволюции своего выражения и тогда продолжают возникать вновь в качестве таких же «доминантных» мутаций.
С другой стороны, мутации с изменчивым выражением и проявлением обнаруживают значительную зависимость не только от индивидуальных свойств генотипа, но и от изменений в факторах внешней среды. Их лабильность проявляется, следовательно, и в индивидуальных реакциях на те или иные частные условия (локальные, временные, сезонные). Такие реакции также могут оказаться частично благоприятными для организма. Они могут иметь хотя бы до известной степени, т. е. в некоторых случаях, положительное значение и могут быть преобразованы в процессе отбора, как показывают эксперименты М. Камшилова (1939б). В таких реакциях мы видим зародыши способности организма к индивидуальным адаптациям как физиологического, так и морфологического характера. Последние обозначаются как приспособительные модификации и являются источником более или менее ясно выраженного полиморфизма данного вида организмов.
Г. СТАБИЛИЗАЦИЯ БЛАГОПРИЯТНЫХ ВЫРАЖЕНИЙ МУТАЦИЙ
Если известные выражения отдельных мутаций имеют хотя бы в некоторых частных условиях существования положительное значение, увеличивая приспособленность организма или его общую жизненную стойкость, то эти выражения становятся предметом естественного отбора. Они преобразуются в процессах комбинирования и усиливаются до достижения известного оптимального уровня. Возможность усиления выражения мутации в гетерозиготном состоянии через ее комбинирование с другими мутациями доказана наблюдениями над дрозофилой. Рецессивная, связанная с полом мутация forked выражается у самца и у гомозиготной самки в укорочении щетинок на голове, груди и скутеллуме. Кроме того, щетинки изогнуты и приподняты. Гетерозиготная самка неотличима от дикой нормы. При ее комбинировании с другой рецессивной мутацией semiforked гетерозиготная самка forked получает, однако, явно промежуточное выражение, т. е. становится «доминантной» (точнее, полудоминантной; D. Lancefield, 1918, по R. Fisher, 1930). Конечно, такой значительный эффект лишь редко достигается комбинированием с одной мутацией, обладающей заметным модифицирующим влиянием. В естественном отборе на жизненную стойкость благоприятных мутаций материал доставляется многими малыми мутациями — модификаторами. Так как этот отбор происходит среди гетерозигот, то повышение положительного выражения мутации в гетерозиготе означает вместе с тем усиление ее доминирования (по меньшей мере в данном ее выражении) . В этом случае возможно быстрое распространение мутации, переход части особей в гомозиготное состояние и отбор на такое же оптимальное выражение в гомозиготе, т. е. на полное доминирование и вхождение ее в состав новой, вполне стабильной нормы. Эволюция доминантности безусловно благоприятной мутации завершается, следовательно, вхождением нового признака или реакции в состав нормы. Вместе с тем в состав нормы включаются в качестве обязательных ее элементов и те модификаторы, которые подавляют неблагоприятные выражения мутации, а также те, которые выправляют и усиливают благоприятные выражения.
Таким образом, постепенно создается определенная генетически сбалансированная система (система геномных корреляций), которая не может быть нарушена без серьезных последствий для нормального формообразования. Устойчивость нормы не нарушается в результате этого постоянного включения в нее новых элементов. Наоборот, постепенное усложнение системы связей все более затрудняет изменение отдельных элементов этой системы. Если же такое изменение наступает, то оно является дезинтегрирующим и по своим последствиям может оказаться летальным. Этот путь эволюции мог бы завести в тупик, если бы не половой процесс и диплоидность (а также регуляторный аппарат), благодаря которым каждая мутация, возникая только в одном геноме, проходит в гетерозиготном состоянии предварительно долгий путь переработки под интегрирующим влиянием стабилизирующей формы естественного отбора.
Однако не всегда дело обстоит так просто. Мы видели, что условная вредность мутации (условная полезность) может привести к более или менее уравновешенному гетероморфизму. К гетероморфизму может привести и частичная вредность (частичная полезность) мутаций. Если благоприятное выражение мутации в результате естественного отбора усиливается в гетерозиготе и достигает оптимального уровня (и доминантности), а неблагоприятное выражение ослабляется и нейтрализуется (т. е. становится рецессивным) также лишь в гетерозиготе, то гомозиготизация данного изменения может вообще оказаться невозможной.
Во всех таких случаях гетерозигота окажется значительно более жизнеспособной, чем гомозигота. Это наблюдается иногда уже на отдельных мутациях. Так, в наблюдениях Леритье и Тессье (Teissier. L'Heritier, 1937) мутация ebony в гомозиготном состоянии не могла конкурировать с дикой нормой и вытеснялась ею, а в гетерозиготном состоянии она поддерживала свое существование на определенном уровне концентрации (20%). Здесь получен, следовательно, экспериментальный «равновесный» полиморфизм искусственной популяции. То же самое происходит, однако, и в природе, если какая-либо мутация оказывается полезной лишь в одном своем выражении. В особенности это касается мутаций с рецессивным летальным или стерильным влиянием. В этом случае гомозиготная мутация оказывается невозможной, а гетерозигота все время расщепляется с образованием гомозиготной нормы и гетерозиготной мутации. Популяция становится равновесно-диморфной.
Если гетерозигота обладает заметными преимуществами перед нормой, то это легко приводит к фиксированию какой-либо другой летали и в гомологичной хромосоме. Тогда приспособленная гетерозиготная форма становится на место нормы и дает начало только таким же гетерозиготным особям (обе гомозиготные формы летальны). Такая система «сбалансированных деталей» способствует, однако, и дальнейшему накоплению неблагоприятных (в гомозиготе) мутаций (Мёллер). Это — крайний случай. В более мягкой форме такие процессы оказываются весьма распространенными.
Примерами высокогетерозиготных популяций, насыщенных деталями, могут послужить виды божьих коровок и, в частности Adalia bipunclata. Гетерозиготность особей обнаруживается как в их разнообразной окраске, так и в физиологических признаках.
Вследствие большой насыщенности популяций Adalia bipunctata деталями, родственное разведение (путем скрещивания братьев с сестрами) оказывается чрезвычайно затрудненным. Оно сопровождается большой гибелью на всех стадиях развития (Я. Я. Лус, 1947)
Отбор идет в природных условиях по гетерозиготам, на базе отбора и фиксирования сбалансированной системы модификаторов, подавляющих вредные выражения мутаций и выдвигающих хотя бы условно благоприятные их выражения. Естественно поэтому, что именно гетерозигота должна, несмотря на ее лабильность, обладать большей приспособленностью и жизнеспособностью. Создаются, таким образом, сложные генетические системы, сбалансированные именно в гетерозиготном состоянии. Полная гомозиготизация таких систем вообще невозможна. Частичная же гомозиготизация, практически достигаемая путем инцухта, ведет к снижению жизнеспособности и к уменьшению плодовитости, как это нередко И наблюдается. Поэтому большая жизнеспособность гетерозиготы и ведет обычно к развитию механизмов, обеспечивающих скрещивание животных и поощряющих перекрестное опыление у растений (R. Fisher, 1930).
Устойчивость гетерозиготы означает, конечно, отсутствие неблагоприятных выражений наличных мутаций в их положительных выражениях. Такое изменение выражения мутаций создается путем комбинирования многих малых мутаций под руководящим влиянием стабилизирующего отбора, т. е. в данном случае отбора в пользу установившейся гетерозиготной нормы. Оно может вести и к тому, что гетерозиготная мутация окажется более устойчивой и жизнеспособной, чем каждая из гомозигот (т. е. как мутация, так и прежняя «норма»). Это явление сверхдоминирования является также результатом действия стабилизирующего отбора по гетерозиготам.
2. ВОЗНИКНОВЕНИЕ ПРИСПОСОБЛЕНИЙ (АДАПТАЦИОГЕНЕЗ)
А. ФИЗИОЛОГИЧЕСКИЕ АДАПТАЦИИ
Обилие и большое значение для жизненной стойкости организма малых физиологических мутаций способствуют, очевидно, тому, что географическая, а частью и экологическая, дифференциация, начинается с небольших различий физиологического характера. Мы уже отмечали, что такие различия в высшей степени характерны не только для невидимых физиологических и для малых мутаций, но и для вполне выраженных морфологических мутаций, являясь обязательным их коррелятом. Это понятно, если мутации по своей природе являются биохимическими изменениями, которые прежде всего ведут к изменениям процессов внутриклеточного обмена веществ и только через это к морфогенетическим преобразованиям. Именно эти элементарные первичные изменения иногда отличаn#1086;рмальный генотип живет при 20о и гибнет при 27° (Banta and Wood, 1927). И отдельные мутации оказываются поэтому в некоторых своих физиологических выражениях благоприятными в обычных условиях культуры. Мутация eversae (D. junebris является при температуре 24–25о более «жизнеспособной» (по проценту вылупления), чем дикая норма (Тимофеев-Ресовский); мутация пурпуровых глаз purple (D. melanogaster) обладает более высокой плодовитостью, а мутация speck связана с большей длительностью жизни. Однако мы знаем, что в некоторых условиях борьбы за существование более низкая плодовитость и даже более короткая жизнь могут приобрести положительное значение.
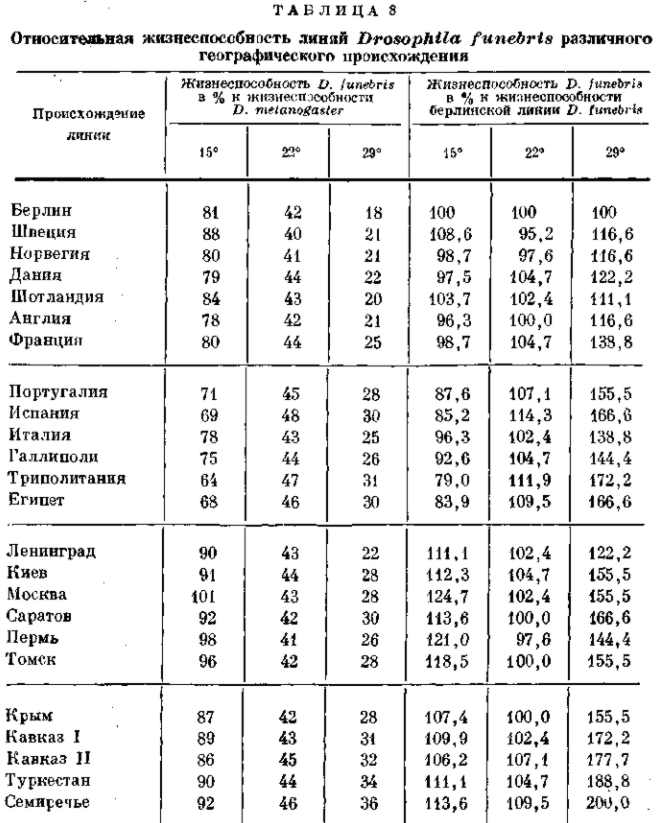
Различия между близкими географическими и экологическими расами и даже между локальными популяциями имеют именно такой характер. Исследования Тимофеева-Ресовского над жизнеспособностью дрозофилы (D. junebris) из различнейших популяций Палеарктики при различных температурах культивирования показали, что юго-западные популяции (из Испании, Италии, Галлиполи, Триполитании и Египта) отличаются большей жизнеспособностью при 29° и меньшей жизнеспособностью при 15° по сравнению с северо-западными популяциями (из окрестностей Берлина, Швеции, Норвегии, Дании, Шотландии, Англии и Франции) ; северо-восточные популяции (из Ленинграда, Киева, Москвы, Саратова, Перми и Томска) оказались более жизнеспособными как при 15°, так и при 29°, а юго-восточные популяции (из Крыма, Кавказа и Средней Азии) оказались также более жизнеспособными при 15°, но в особенности отличались высокой жизнеспособностью при высоких температурах (29°) (табл. 8). Эти различия находятся в полном согласии с умеренным морским климатом северо-запада, с теплым и мягким климатом юго-запада, с прохладным континентальным климатом северо-востока и с резко континентальным климатом юго-востока. Континентальные популяции отличаются именно высокой стойкостью по отношению к крайним температурам — как низким, так и высоким.
То же самое было найдено Добжанским в отношении североамериканских популяций дрозофилы (D. pseudoobscura). В этом случае изучалась их плодовитость при разных температурах. Оптимальная для плодовитости температура оказалась для более северной и западной расы В ниже, чем для юго-западной расы А К Обычно D. pseudoobscura не выносит температур выше 27,5°. Однако в штате Аризона в окрестностях Юмы были найдены две линии, относящиеся к расе Л, которые удалось культивировать в лаборатории при 28,5–39°. Местность, в которой они были найдены, отличается исключительно жарким летом. Морфологически эти расы неразличимы. У жужелиц Carabus nemoralis географические формы ясно различаются по предпочитаемой температуре.
Чувствительность к разным температурам лежит нередко в основе расовых различий у рыб. «Озимые» и «яровые» расы проходных и полуироходных рыб начинают свои миграции именно при разных температурах воды (Л. Берг, 1934); яровые поднимаются в реки при более высокой температуре и не идут так далеко вверх по течению. По Ильину (1926), географические расы горностаевых кроликов обладают весьма различным температурным порогом образования черного пигмента (сравнивались московская, польская и две германские расы, из них московская обладает наиболее низким, а германские наиболее высоким верхним порогом пигментооб-разования). У многих млекопитающих обнаруживаются расовые различия в предпочитаемых температурах. В экспериментальных условиях, с наличием температурного градиента, серая мышь предпочитала 37,3°, а альбиносы — 34,6° (Гортер). Соответственно и температура их тела различна (36,96° и 33,87°).
Установлены и многие другие расовые различия (а также различия между разными линиями): стойкость к ядам, к паразитарным заболеваниям, иммунитет, морозоустойчивость и засухоустойчивость растений и многие др. Как правило, все эти различия оказываются приспособительными в данных жизненных условиях. В расовых различиях индивидуального развития растений большую роль играют условия влажности и температуры (яровые и озимые расы с дальнейшими различиями) и длина светового дня. Те же факторы играют большую роль в половом цикле птиц и могут быть основой расовых различий. Расхождение во времени цветения у растений и времени полового размножения у животных покоится иногда на очень небольших различиях в физиологических реакциях на изменения во внешних факторах, и, однако, они оказываются часто приспособительными и имеют огромное значение в расо- и видообразовании, так как ведут к половой изоляции новых от исходных форм. Мы в этой связи упоминали о различиях во времени размножения близких рас бабочек (например, дубового шелкопряда), различных рас рыб (сельдей и проходных рыб) и амфибий. То же самое касается, однако, птиц, млекопитающих и очень многих других животных. У млекопитающих имеются нередко большие расовые различия в их поведении, в общей активности и в суточном поведении (дневная, ночная и т. д.). Такие физиологические различия имеют также приспособительное значение; они связаны с местными условиями добывания пищи и защиты от хищников (см. работы Калабухова, Наумова и др.). Все это — определенные экологические различия, которые, как видно, нередко имеют в первую очередь физиологический характер. Это касается и расхождения в питании животных, в формах заботы о потомстве и мн. др.
Равнинные и горные формы живородящей ящерицы (по Рихтеру, 1933), а также лесной мыши (по Калабухову, 1937) обладают наследственными различиями в количестве эритроцитов и в содержании гемоглобина в крови. По Н. Калабухову (1939), северные формы лесной (Apodemus sylvaticus) и желтогорлой (A. flavicollis) мыши отличаются от южных более интенсивным обменом (потреблением кислорода), большей активностью и более низкой предпочитаемой температурой. Близкие и морфологически не отличимые, локальные формы улиток Lymnaea columella из двух разных водоемов (в Пенсильвании, по Бейли, 1939) отличаются по длительности жизни, плодовитости и скорости роста. Гусеницы двух морфологически не различимых рас бабочки Hyponomeuta padella отличаются тем, что одна из них охотнее откладывает яйца на яблоне (90,2%), а другая —на листьях шиповника (79,3%). Очень много фактов такого рода собрано в сводке Калабухова (1941а). Для экологических форм растений физиологические различия не менее характерны, чем для животных. В особенности это касается таких признаков, как засухоустойчивость, устойчивость по отношению к инфекциям и грибным заболеваниям. Экотипы Турессона различаются также по своей физиологической характеристике. Последняя тесно связывается и с морфологическими различиями (см. сводки Розановой, 1940, и Синской, 1948).
Б. ПРИСПОСОБИТЕЛЬНАЯ ОКРАСКА
Пигментация животных по своему развитию и изменчивости приближается к физиологическим признакам. Различия в пигментации также очень тесно коррелированы с физиологическими различиями. В особенности это касается альбиносов, которые обычно отличаются более вялым обменом (хотя белые крысы развиваются, растут и достигают половой зрелости раньше серых) и меньшей активностью. Это верно, однако, и по отношению к меланистиче-ским формам, которые также, по-видимому, обычно обладают и физиологическими различиями. Нас интересует здесь, однако, не этот вопрос, а приспособительное значение расовых различий в окраске. И здесь относительная простота таких изменений ведет к тому, что самые различные уклонения могут оказаться благоприятными в тех или иных частных условиях. Большая или меньшая интенсивность окраски может иметь защитное значение в соответствующих условиях общего фона и освещения.
Распространение меланистической формы хомяка приурочено к влажньщ районам лесостепи на Украине и в Башкирии (С. Гершензон, 1946). Быть может, здесь темная окраска дает ему большую защиту, чем в открытой степи. Однако С. Гершензону.не удалось установить фактов различной истребляемости обеих форм хищниками. Расселение меланистической формы лимитируется зимней гибелью, и это заставляет думать о значении каких-то физиологических коррелятов (в осенних популяциях процент меланистов возрастает либо в результате большего выживания в течение лета, либо в силу большей плодовитости; в весенних популяциях процент меланистов ниже вследствие большей зимней смертности). Здесь защитное значение окраски не доказано. Однако вариации в окраске весьма распространены и во многих случаях, несомненно, приобретают защитное значение (а в других случаях — значение распознавательных отметин, как, например, у многих пауков и у птиц).
Так, например, расовые различия в окраске шерсти североамериканской мыши Peromyscus polionotus, живущей во Флориде, имеют, по Сёмнеру, защитное значение. Обычная серая окраска P. polionotus polionotus под цвет почвы полей заменяется на песчаном побережье очень светлой окраской P. polionotus albifrons. На маленьком полуострове имеется третий подвид P. polionotus leucocephalis, еще более светлый, вполне подходящий к белому цвету песчаной поверхности полуострова. Эти различия оказались наследственными. То же самое было найдено относительно различий в окраске ряда других грызунов, в частности — рода Perogna-thus, которые, по Бенсону, имеют также приспособительный под цвет почвы характер. На песчаной почве распространены светлоокрашенные, иногда почти белые локальные расы этих грызунов, а на темных лавах — темноокрашенные географические или экологические расы.
Наконец, в известных исследованиях Гаррисона (Harrison, 1920) был показан механизм самого возникновения различий в окраске в двух популяциях бабочек ОрогаЫа autumnata, возникших из одной сплошной популяции при разделении леса широкой просекой. В той части леса, где сосна была замещена березой, естественный отбор (преобладающее истребление птицами более темных особей) привел к значительному посветлению популяции бабочек. Различия в окраске географических и экологических рас имеют, как сказано, наследственный характер и покоятся на комбинациях мутаций, которые иногда производят впечатление очень простых различий в отдельных факторах (например, различные формы богомолов или различные мимикрирующие формы самок Papilio polytes и Р, darclanus).
В. ИНДИФФЕРЕНТНЫЕ МОРФОЛОГИЧЕСКИЕ РАЗЛИЧИЯ
Систематики постоянно подчеркивают, что расовые и видовые различия имеют главным образом индифферентный характер и не могут рассматриваться как приспособления. К таким утверждениям следует всегда относиться с осторожностью, так как мы в огромном большинстве случаев не знаем экологических и особенно биоценотических соотношений с такой полнотой, чтобы иметь правильное суждение о физиологическом и биологическом значении тех или иных признаков. Тем не менее, если бы это было и так, в этом нельзя усмотреть затруднений для теории естественного отбора, против которой обычно такие аргументы выдвигаются.
Прежде всего отметим, что положительная роль естественного отбора основывается всегда на обратном процессе — избирательной элиминации менее приспособленных особей. Поэтому вместе с накоплением благоприятных изменений идет всегда и накопление безразличных мутаций, малых мутаций, а также и условно вредных и частично вредных мутаций. Вредные выражения мутаций нейтрализуются в процессе стабилизирующего отбора модификаторов, а безразличные выражения могут остаться в качестве признаков рас и даже видов. Они могут быть использованы и в качестве распознавательных отметин (приобретают тогда положительное значение в процессе полового размножения, в уходе за потомством и в стадной жизни).
Даже такой совершенно безразличный морфологический признак, как правое или левое вращение улиток, оказался связанным с физиологическими различиями (Гаузе, Смарагдова, 1939). Улитки (Fruticicola lantzi) с левозавитыми раковинами в лабораторных условиях быстро падали в весе, по всей вероятности, вследствие более интенсивного обмена. Между тем, в Казахстане в некоторых местах (в окрестностях Алма-Ата) левозавитые раковины составляют громадное большинство популяций (в других местностях полностью господствуют правозавитые улитки).
Близкие виды постоянно отличаются по плодовитости, скорости созревания, темпу размножения, по времени спаривания и размножения, по питанию, характеру активности и ее распределению в дневное и ночное время. Исследование всегда показывает связь расовых, подвидовых и видовых различий с особенностями местообитания и жизни в определенной биоценотической обстановке. Неизменно эти различия оказываются приспособительными.
Современные исследования экологов дают огромный материал для физиологической оценки различий между близкими видами, подвидами и даже отдельными популяциями одного и того же вида или подвида. В этом отношении большое значение имеют прекрасные работы Калабухова по изучению близких форм грызунов, которые ясно показывают адаптивность этих различий. Весьма возможно, что во многих случаях физиологическая адаптация действительно предшествует морфологической (Промптов, 1933). Однако вряд ли это составляет общее правило. Детальные исследования морфологических признаков вскрывают и здесь адаптивность подвидовых и расовых различий. Напомню результаты исследований окраски экологических рас и подвидов различных грызунов, проведенных рядом американских авторов (Sumner, Dice, Benson и др.). Эти исследования показали защитный характер окраски даже отдельных популяций — от почти белой на песчаной почве до почти черной на темных лавах. Сошлюсь также на исследования видов и подвидов рода Dipodomys (Grinell I., 1922), показавшие приспособленность к бегу скачками в определенных условиях открытых степей (более длинные конечности) или кустарных зарослей (конечности короче и толще), а также на исследования видов и подвидов рода Passerella (Linsdale I., 1928) в отношении развития скелетных частей крыла, плечевого пояса и грудины в связи с областью гнездования и длиной пути их перелетов на юг (развитие этих частей тем выше, чем длиннее пути миграции).
Во всех тех случаях, когда экологи производят детальное исследование, вскрывается адаптивный характер не только физиологических, но и морфологических видовых различий. Почему же все-таки систематики так усиленно подчеркивают «индифферентный» характер видовых различий? Это объясняется именно тем, что адаптивные видовые различия неудобны для систематика — они не всегда видны на консервированном материале (физиологические различия) или требуют более сложного анатомического исследования («организационные» различия), и притом обычно — ввиду трансгрессивного их характера — большого материала и биометрической его обработки. Систематики идут по этому последнему пути; однако и этот путь дает возможность установить адаптивный характер различий лишь при наличии исчерпывающих наблюдений над жизнью данного организма в его естественной обстановке. Следует учесть и еще одно обстоятельство, когда сравнивают адаптивность видовых различий с адаптивностью признаков более крупных таксономических подразделений.
Конкретным объектом эволюции являются всегда только виды с их видовыми различиями. Виды превращаются в другие виды, дифференцируются и дают начало более или менее многочисленным новым видам. При всем этом виды всегда остаются видами. Признаки организации меняются в связи с изменением соотношений с элементами внешней среды, и какие из этих изменений будут иметь ограниченный характер «видовых» различий, а какие в дальнейшем процессе эволюции приобретут более общий характер родовых, порядковых или классовых различий,— заранее предсказать невозможно. Только ретроспективно -мы можем сказать, что такие-то различия, возникнув когда-то в качестве видовых, приобрели затем значение признаков, характеризующих семейство или другое крупное подразделение. Одни признаки имеют кратковременное существование — они связаны с ограниченными условиями жизни данного вида и исчезают вместе с его преобразованием в другой вид, живущий при иных условиях. Другие признаки имеют более длительное существование — они сохраняют свое значение и при некотором изменении внешней обстановки и, следовательно, при распаде вида на целую группу видов. Естественно, что более длительное время сохраняют свое значение признаки явно адаптивного характера. Признаки наиболее общие именно потому и входят в характеристику крупных таксономических подразделений, что их адаптивный характер не теряется с изменением локальных — географических или экологических — условий существования (хотя в деталях они, конечно, изменяются). Маловажность других различий, которые могут иметь весьма ограниченное значение в данной местности, в данных экологических условиях или для данной популяции (например, распознавательные отметины), является причиной их легкой изменяемости и сравнительно кратковременного существования. Маловажность признаков затрудняет вскрытие их адаптивности (без специального исследования в естественной обстановке), а с другой стороны, она ограничивает их значение пределами низших систематических подразделений. Систематики, подчеркивающие широкий, приспособительный характер признаков больших подразделений и нейтральный характер видовых различий, попадают, следовательно, в логический круг. Различия ограниченного значения, т. е. «нейтральные» признаки, потому и избираются для видовой диагностики, что они резко изменяются вместе с изменением самого вида. Признаки гораздо более важные — «организационные» — меняются медленнее, и их преобразования, нередко количественные, гораздо менее удобны для систематика. Мы не раз подчеркивали ведущее значение центральной нервной системы в эволюции высших позвоночных. Определяя характерное поведение данного вида, подвида и даже экологической расы животных, видовые различия в строении центральной нервной системы имеют, несомненно, адаптивное значение. Вряд ли, однако, было бы удобно класть в основу видовой диагностики специфику микроскопической архитектоники головного мозга.
Г. МОРФОЛОГИЧЕСКИЕ АДАПТАЦИИ
Морфологические выражения мутаций наиболее изучены. Однако как раз здесь трудно установить существование положительных изменений, которые могли бы иметь значение адаптации. Приходится признать, что морфологические признаки, будучи конечными звеньями в цепи биохимических и физиологических реакций и основанные всегда на сложных онтогенетических взаимозависимостях, чаще всего испытывают очень грубые изменения. Поэтому морфологические адаптации могут строиться только на очень малых мутациях, не имеющих значительного деструктивного характера.
Нетрудно, конечно, понять явления редукции органов, потерявших свое значение, так как большинство мутаций связано именно с явлениями недоразвития. Такие адаптивные преобразования, как рудиментация крыльев у островных насекомых, редукция глаз у пещерных и роющих животных или редукция конечностей у змееобразных позвоночных, настолько ясны, что на этом можно не останавливаться (Шмальгаузен, 1938а, 1942).
Не представляет особых трудностей и вопрос о количественных изменениях, лежащих в основе прогрессивного развития органов. Форма, число и длина щетинок у дрозофилы, форма и длина конечностей и антенн, число омматидий в глазах, число яйцевых трубок и яйцевых камер — все эти величины меняются во многих весьма обычных мутациях. В основе новообразований могут также лежать и чисто количественные изменения. К числу весьма распространенных мутаций относятся всевозможные удвоения, ведущие к увеличению числа общегомологичных органов — удвоение щетинок у дрозофилы, удвоение конечностей. У позвоночных бывают частичные удвоения конечностей (полидактилия у кур, голубей; также у млекопитающих), увеличение числа сегментов тела и т. п. Такие изменения лежат, очевидно, в основе явлений «полимеризации», которые могут вести к новым, более разнообразным дифференцировкам между общегомологичными органами. В процессе эволюции такие изменения играют, очевидно, немалую роль.
Гораздо труднее проследить за начальным возникновением качественно новых адаптации. Фактических наблюдений этого рода имеется пока еще очень мало. Наиболее вероятным представляется мне предположение, что такие совершенно новые адаптации возникают именно на базе тех «нейтральных» различий, которые в большом числе накапливаются в составе «резерва» внутривидовой изменчивости и в процессах перекомбинирования мутаций непрерывно меняют свое выражение. При изменениях внешней среды или при миграции самого организма могут сложиться и новые, более благоприятные соотношения между организмом и средой.
Изменения в жилковании крыла принадлежат к наиболее распространенным выражениям разнообразных мутаций дрозофилы. Вместе с тем тип жилкования представляет важный и довольно устойчивый систематический признак, характеризующий целые группы насекомых и, очевидно, не лишенный приспособительного значения (он определяет упругие свойства крыла как летательного органа). Некоторые крыловые мутации все же распространяются в популяции и могут послужить материалом для приспособительных преобразований конструкции крыльев. У видов рода Andrena (Hymenoptera) встречается мутация second cubital crossvein. У A. albicans она наблюдается очень редко, у А. ргаесох и особенно у A. vaga — несколько чаще. У A. serica эта мутация довольно обычна (около 8% особей), а у A. argentata встречается еще чаще (около 20% особей). Очевидно, эта мутация приобретает уже какое-то положительное значение. Наконец, у A. neglecta это изменение стало нормальным признаком вида (Zimmermann К., 1933). Качественный характер имеет и мутация simplex полевки Microtus arvalis, выражающаяся в изменении строения коренных зубов. Эта мутация распространяется в Шлезвиг-Гольштинии, достигая там значительной концентрации — выше 85% особей, что указывает на ее положительное значение в данных условиях.
Если мы говорили раньше о частично вредных или обезвреженных морфологических коррелятах физиологических мутаций и возникающих на их базе физиологических адаптации, то теперь мы должны подчеркнуть, что и морфологические изменения обладают не только физиологическим выражением, но и своими коррелятивными связями. Если добавочные физиологические и морфологические (плейотропные) выражения морфологических мутаций не слишком неблагоприятны (снижение «жизнеспособности» и «плодовитости»), то они нейтрализуются в процессе отбора наиболее благоприятных комбинаций. Тем самым они теряют свои второстепенные выражения. Однако всякий морфологический признак играет какую-то роль в жизни организма. Поэтому, входя в состав приспособленной нормы, морфологическое изменение всегда приобретает если не физиологическое, то биологическое или экологическое значение.
Физиологическое выражение морфологических адаптации состоит в характерных для них жизненных функциях организма. Так, даже изменение величины тела может иметь физиологическое значение. Увеличение размеров тела связано всегда с более экономным обменом веществ, так как продукция энергии определяется объемом живого вещества или активных тканей организма, а трата энергии — поверхностью организма. Это касается всех животных, а не только гомойотермных. Пойкило-термные животные лишь непрерывно растрачивают продуцируемую тепловую энергию, отдавая ее в окружающую среду, а го-мойотермные животные частично сберегают, используют и поддерживают температуру на оптимальном уровне. Поэтому «правило Бергмана» о возрастании величины тела (близких групп животных) при переходе от теплого климата к более холодному частично распространяется и на «холоднокровных» животных. Однако значительная интенсивность обмена (и жизнедеятельности) у птиц и у млекопитающих естественно ставит вопрос о потере энергии через теплоотдачу более остро, чем у других животных. Поэтому правило Бергмана имеет здесь более всеобщий характер. Большую величину тела у птиц и млекопитающих холодных стран мы должны считать морфологическим признаком большого физиологического значения. То же самое относится к таким характеристикам, как величина ушей у некоторых млекопитающих и длина хвоста у мышей. Во многих случаях они имеют прямое отношение к терморегуляции (как части тела с менее развитым волосяным покровом и богато развитой сетью кожных кровеносных сосудов). Большие уши южных лисиц способствуют теплоотдаче и предохраняют тело от перегревания, а малые уши песца, наоборот, сокращают возможную потерю тепла. Длинные хвосты южных мышей и короткие хвосты северных имеют такое же физиологическое значение.
Чисто морфологические различия в числе, длине и толщине волос у млекопитающих играют такую же роль в регулировании теплоотдачи. Эти изменения исключительно просты, и вместе с тем их адаптивность совершенно ясна. Приводим таблицу 9, показывающую различия в длине и толщине волос у северных и южных млекопитающих.
Интересную попытку проследить возникновение морфологических адаптации определенного функционального значения сделал Н. В. Дубовский (1941) в своем сравнительном и экспериментальном исследовании органов движения и прикрепления у ракушковых рачков (Ostracoda). Эти рачки плавают при помощи двух пар антенн, снабженных длинными гребными щетинками, и ползают, опираясь на вторую пару ножек, снабженных конечными коготками (у морских форм для хождения используются три пары ножек).
Формы, исключительно ползающие, характеризуются укорочением гребных щетинок первой пары антенн, рудиментацией или даже отсутствием щетинок второй пары антенн; укороченными, утолщенными и более мускулистыми конечностями, особенно у форм, зарывающихся в песчаный грунт. В пределах многих родов пресноводных плавающих остракод имеются отдельные виды, приспособленные исключительно для ползания. Эти последние характерны для текучих вод и, очевидно, произошли от исходных форм стоячих водоемов.
Для экспериментального исследования избран типичный представитель речных Ostracoda (Волги и Сев. Донца) — Cypria curvifurcata, У этого вида установлено существование различий между локальными формами, заселяющими места реки с песчаным дном и более сильным течением, и формами, заселяющими участки с заиленным дном и медленным течением. Биометрическая обработка материала, полученного в результате многочисленных измерений, показала существование вполне достоверных различий по очень многим показателям. В общем формы, живущие на песчаном дне участков рек с более сильным течением, имеют конечности и их придатки (коготки и щетинки) сравнительно короткие, толстые, сильнее хитинизированные, очевидно, приспособленные к передвижению на крупнозернистом песке, к закапыванию в нем и к укреплению и противостоянию более сильному течению. Формы с заиленных мест рек со слабым течением характеризуются сильным развитием придатков конечностей (длинные и тонкие коготки и щетинки) и большей длиной самих конечностей. Длинные конечности характерны и для лазающих по растениям представителей Cypridae из стоячих пресных водоемов. Сравнительно-морфологический анализ морских Ostracoda (сем. Cytheridae) также показывает, что длинные конечности с тонкими придатками имеются у видов, живущих на более зыбком илистом грунте. Эксперименты поставлены следующим образом. Особи, взятые с заиленных мест реки, помещались в деревянный лоток вместе с некоторым количеством грунта и подвергались действию тока воды, который смывал животных в подставленный сачок. Оставшиеся животные смывались затем более сильной струей воды в другой сачок. Смытые вначале животные первого сачка сравнивались с оставшимися животными из второго сачка (после фиксирования, измерения и биометрической обработки полученных величин). По 12 признакам получены статистически достоверные различия между смытыми и несмытыми особями. Именно у несмытых особей оказались более короткие коготки второй антенны, более короткие дистальный членик и коготок первой ножки и более короткие основание и коготки фурки. По этим признакам несмытые особи приближаются к локальным формам, характерным для мест с более быстрым течением. Это показывает приспособительный характер изменений и эффективность отбора по перечисленным признакам в экспериментальных условиях, причем направление этого отбора совпадает с направлением изменений в естественных биотопах.
ТАБЛИЦА 9. Длина (в мм) и толщина (в мк) волос географических рас, обитающих в разных климатических поясах (по Rensch, 1929)
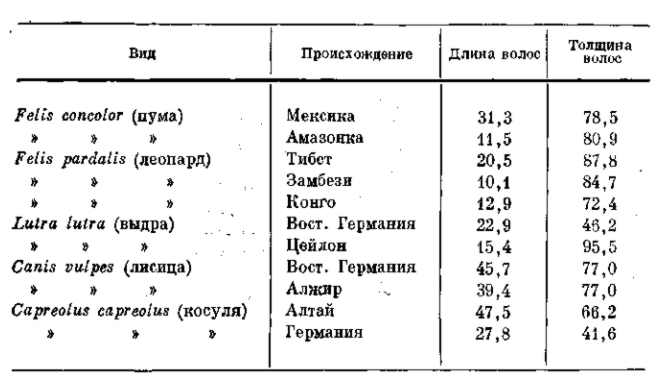
При изучении морских Ostracoda из сем. Cytheridae найдены подобные же различия между прибрежными формами, живущими в зоне прибоя, цепляющимися за водоросли и обладающими наиболее короткими члениками конечностей и коготками, формами, ползающими на песчаном грунте, и, наконец, формами, живущими на илистом дне. Эти последние обладают наиболее длинными конечностями, щетинками и коготками, приспособленными не столько для цеплянья и лазанья, сколько для того, чтобы при помощи увеличенной поверхности держаться на зыбком иле, а также зарываться в него.
Экологическое значение морфологических изменений определяется именно положением организма во внешней среде и, в особенности, его соотношениями с другими организмами. Таковы уже упомянутые явления защитной окраски, таковы же и различные другие средства пассивной (а также и активной) защиты. Хитиновые, роговые и костные наружные панцири с шипами и иглами могли развиваться на базе самых незначительных и очень простых изменений адаптивного характера (в смысле относительной эффективной защиты от хищников или паразитов).
Многие морфологические приспособления были бы совершенно непонятны, если бы мы не знали их экологического и, в частности, биоценотического значения. Таковы взаимные приспособления организмов. Как раз в этих случаях нелегко себе представить адаптивность ничтожных различий во взаимных приспособлениях различных форм одного вида или совершенно различных организмов. Трудность увеличивается здесь именно необходимостью взаимного согласования этих изменений. В особенности это касается половых различий. И все же половые части многих насекомых в высшей степени пластичны и всегда хорошо согласовываются в обоих полах. Приходится и здесь допустить, что естественный отбор имеет дело только с очень малыми изменениями, которые не вносят существенных нарушений в половые взаимоотношения.
Не столь очевидно строги требования взаимного приспособления цветов и насекомых-опылителей, поскольку узкая специализация наблюдается здесь довольно редко. Нетрудно понять, что даже небольшое удлинение хоботка пчелы может иметь приспособительное значение, позволяя использовать для добычи меда также растения с более глубокими нектарниками К Четыре вида небольших жуков рода Diharda незначительно отличаются друг от друга по своим размерам и окраске. Трудно было бы признать адаптивное значение этих ничтожных различий, если бы не было известно, что они относятся к мирмекофилам и, живя в муравейниках, соответствуют по размерам и окраске четырем различным видам муравьев, среди которых они живут.
Если существуют известные трудности для изучения возникновения новых приспособлений в природной обстановке, то это восполняется рядом очень интересных наблюдений над нашими культурами. Возникновение морфологических различий экологического характера можно прекрасно проследить на истории животных и растений в искусственных условиях и, в особенности, на истории вредителей сельского хозяйства и сорняков полевых культур, которые показывают приспособление организмов к новым условиям, создаваемым человеком.
Уже исследования Веттщтейна над влиянием покоса на растительность альпийских лугов показали наличие значительных изменений характера «приспособления» к деятельности человека. Подобные исследования были затем предприняты Н. В. Цингером, который показал, что, например, горный вид погремка (Alectorolophus montanus), цветущий на нетронутых альпийских лугах в течение всего лета, дал на укосных лугах начало рано зацветающему летнему виду (A. aestivalis). В условиях двух покосов этот вид разбился на два — раннеспелый весенний вид (A. vernalis) и позднеспелый летний (A. polycladus). Еще интереснее, что в условиях посевов ржи летняя форма преобразовалась в особую форму, приспособившуюся к этим посевам (A. apterus). У этой формы коробочки не раскрываются и семена сами не осыпаются (они требуют обмолота!). Крылья, бывшие на семенах исходных форм, редуцировались, и поэтому семена не отвеиваются. Они остаются в урожае ржи. Таким образом этот вид погремка превратился в сорняк. Эволюция сорняка, однако, еще не вполне закончена. У этой формы (A. apterus) еще сохранились признаки, указывающие на ее происхождение: иногда попадаются коробочки с щелью, через которую семена еще могут высыпаться; среди семян часто встречаются, кроме бескрылых, также и семена с узкой оторочкой—рудиментом крыльев (52% семян) и в небольшом числе (12%)—с заметными или даже (1,5%) хорошо развитыми крыльями.
В условиях систематического покоса происходит также отбор низкорослых особей растений — так, например, устанавливается низкорослая форма лебеды, не уничтожаемая при кошении. Интересны и другие исследования Н. Цингера над происхождением сорняков и в особенности над растениями, обычно засоряющими посевы льна. Льняной рыжик (Camelina linicola) близок и, очевидно, произошел от ярового рыжика (С. glabrata), отличаясь от него прежде всего более крупными семенами, которые по весу приближаются к семенам льна. То же самое касается льняной гречихи {Polygonum linicola), происходящей от дикой P. lapathifoliurn, льняной торицы (Spergula linicola), происходящей от обыкновенной S. vulgaris, льняного плевела (Lolium remotum), происходящего от обыкновенного сорняка яровых посевов L. temulentum. Происхождение этих форм связано с бессознательным отбором, так как при просеивании семян льна всегда оставались самые тяжелые семена этих растений, наиболее приближающиеся по весу к семенам льна. Вес семян был ведущим морфологическим признаком, по которому шло «приспособление» новых видов-сорняков к условиям жизни и размножения в посевах льна.
Новейшие данные, касающиеся происхождения культурных злаков, показывают, что рожь и овес вошли, очевидно, в человеческую культуру первоначально как сорняки посевов пшеницы и ячменя. По мере продвижения культуры на север они все более засоряли посевы, но вместе с тем приобретали все большее значение, сначала как непроизвольная и безвредная примесь, а затем как самостоятельный объект культуры.
Наконец, и вообще история культивируемых растений и домашних животных дает картину очень постепенного физиологического и морфологического «приспособления» этих организмов к своеобразным экологическим условиям, создаваемым человеком. Далеко не все эти изменения направляются человеком. Последнего интересуют всегда лишь очень немногие свойства. Остальные сопутствующие изменения могут иметь характер обычного «приспособления», либо иногда (чаще, чем в природной обстановке) характер действительно вполне «индифферентных» признаков.
3. ВОЗНИКНОВЕНИЕ АДАПТИВНЫХ ФОРМООБРАЗОВАТЕЛЬНЫХ РЕАКЦИЙ
Хотя и рассмотренные физиологические и морфологические приспособления развиваются на основе измененной нормы реакций организма, вопрос о возникновении способности к адаптивным реакциям организма все же гораздо сложнее, чем рассмотренные вопросы, так как здесь речь идет не об одной типичной мор-фогенетической реакции, а о возможности целого ряда таких реакций, сохраняющих свою адаптивность при изменении факторов внешней среды и изменении положения организма в этой среде.
Вместе с тем этот вопрос имеет большое общее значение, так как индивидуальная приспособляемость организма создает для него особое положение весьма гибкой устойчивости, т. е. определенной «лабильности», позволяющей ему не только переживать периоды значительных и резких изменений в факторах среды, но и активно переходить из одной среды в другую и даже перестраивать свою организацию. Поэтому проблема развития системы адаптивных реакций организма имеет большое значение и для понимания закономерностей эволюционного процесса.
Кроме того, она относится к числу крайне мало разработанных вопросов эволюционной теории, так как ламаркисты исходили в своих построениях из фактов индивидуальной приспособляемости как заранее данной предпосылки, не вникая в вопрос об ее возникновении. Неодарвинисты же не придавали ему значения, так как предполагалось, что результаты индивидуального приспособления, не будучи наследственными, не имеют значения в процессе эволюции. Мы здесь рассматриваем эти вопросы только в отношении морфогенетических реакций.
А. ЭЛЕМЕНТАРНЫЕ ЗАВИСИМОСТИ РЕАКЦИИ И ИХ ПРЕОБРАЗОВАНИЕ
а. Зависимое развитие морфозов. Элементарные формы реагирования мы усматриваем в тех изменениях индивидуального формообразования, которые наступают в связи с изменениями в факторах внешней среды и не имеют еще под собой заметной исторической базы, а могут рассматриваться как новые реакции. Такие изменения мы называем морфозами, Если они оказываются сходными с известными мутациями, то их называют также фенокопия-ми. Морфозы вызываются чаще всего действием необычных факторов — лучей Рентгена, химических соединений или обычными факторами при необычайной их интенсивности, например, крайне высокими или крайне низкими температурами. В таких морфозах нет ничего приспособительного — это такие же нарушения развития, как и видимые мутации. Большого интереса для нас они здесь не имеют. В разбираемом вопросе генезиса адаптивных модификаций имеют значение главным образом лишь такие реакции или морфозы, которые связаны с обычными в нормальной среде изменениями внешних факторов. Такие морфозы отличаются не столь резким выражением, они не имеют такого «патологического» характера, как обычные рентгено-хемо- или термоморфозы. Однако частично именно поэтому они представляют особый интерес.
Известна зависимость выражения многих мутаций от изменений внешних факторов. Некоторые мутации, как, например, мутация безглазия (eyeless) у дрозофилы или мутация уродливого брюшка (Abnormal abdomen) получают свое полное выражение лишь при развитии личинок в свежей питательной среде; на подсохшем корму развиваются мухи, приближающиеся по своему фенотипу к дикой норме. Другие мутации зависят в своем выражении от температурных условий. Мутация удвоения конечностей (reduplicated) дрозофилы получает свое типичное выражение лишь при низкой температуре. При обычной температуре культивирования личинок ее выражение исчезает. Интересно, что для близких мутаций такие зависимости могут иметь прямо противоположный характер. Так, мутация рудиментарных крыльев (vestigial) дрозофилы получает свое наиболее ясное выражение при низких температур культивирования; по мере повышения этой температуры размеры крыльев увеличиваются, и при 32° у известного числа мух развиваются почти нормальные крылья (рис. 22). С другой стороны, аллеломорфная (по отношению к vestigial) мутация pennant, обладающая также редуцированными крыльями, получает свое полное выражение только при высокой -температуре культивирования. По мере понижения этой температуры в пределах от 30 до 16о размеры крыльев увеличиваются и достигают нормы (рис. 23). У двух аллеломорфных мутаций температурная реакция тканей одного и того же зачатка органа изменяется в противоположных направлениях. Нормальный же аллеломорф этих мутаций отличается максимальной устойчивостью и дает лишь слабую температурную реакцию. Интересно, что оба мутантных аллеломорфа вместе (компаунд vestigial X pennant) дают закономерную смену направления реакции: при повышении температуры в пределах от 16 до 22° культивирования величина крыльев уменьшается (как у pennant), при повышении температуры от 22 до 26° размеры крыльев остаются неизменными {морфогенез устойчив), а при дальнейшем поднятии температуры от 26 до 32е крылья увеличиваются (как у vestigial; см. рис. 24, табл. 10. Это показывает, что характер реакции зависит не от отдельного «гена», а от их комбинации и от всего генотипа в целом. Мы уже обращали внимание на то, что наиболее изменчивым выражением обладают многие рецессивные мутации в гомозиготе (например, eyeless, vestigial, pennant, reduplicated) и полудоминантные мутации в гетерозиготе (напр., Bar, Abnormal abdomen). Это касается как их чувствительности к изменениям в факторах внешней среды, так и легкой изменяемости реакций и форм реагирования в зависимости от незначительных изменений генотипа (при комбинировании малых мутаций — модификаторов). Поэтому и форма реагирования может быть изменена не только путем внесения второй мутации (как в примере vestigial X pennant), но и в результате искусственного (а следовательно — и естественного) отбора малых изменений, зависящих от небольших различий в генотипе (рис. 25).
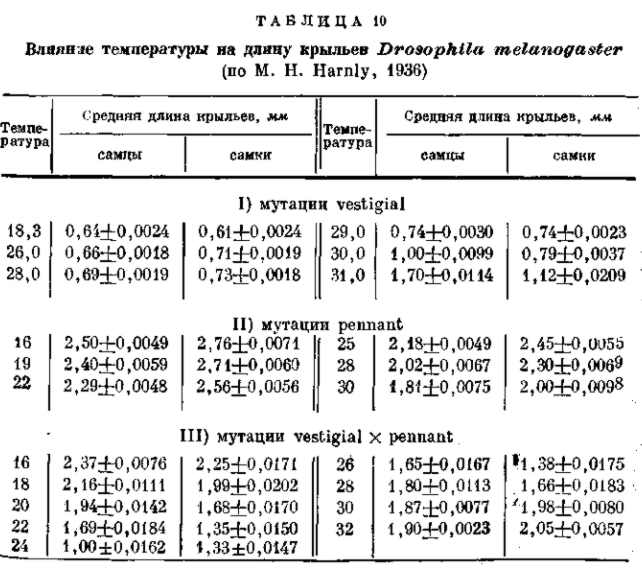
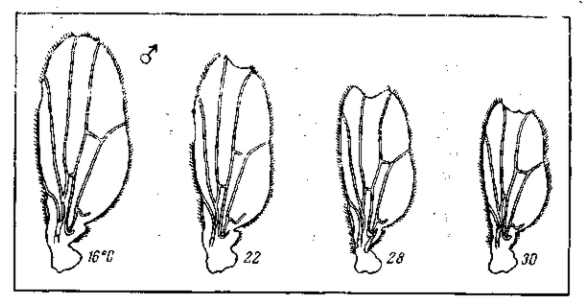
Рис. 23. Зависимость формы и величины крыльев самцов мутации pennant (Drosophila melanogaster) от температуры. Эта мутация показывает реакцию, обратную по сравнению с ее аллеломорфом vestigial. В этом случае при низкой температуре 16° С около 13% особей имеют совершенно нормальные крылья. По М. Г. и М. Л. Харнли, 1935
Экспериментов, показывающих эффективность искусственного отбора на усиление или ослабление выражения известных мутаций, было проведено очень много. Нас интересует здесь, однако, не просто возможность изменения реакции и ее результата — выражения мутации (об этом мы уже говорили), а возможность изменения характера зависимости этого выражения от факторов внешней среды (рис. 26).
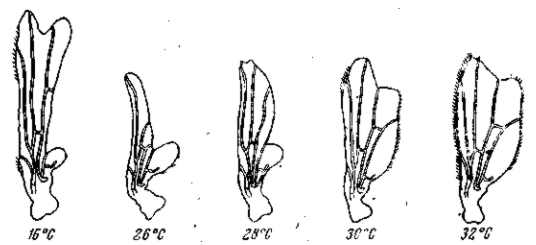
Рис. 24. Зависимость формы и величины крыльев гетерозиготы vestigial X pennant от температуры. Реакция меняет свое направление. При повышении температуры от 16 до 220 крылья уменьшаются; при 22–26° их средние размеры сохраняются постоянными; при дальнейшем повышении температуры крылья вновь увеличиваются. По М- Г. и М. JI. Хари, 1936.
В этом направлении был поставлен ряд интересных экспериментальных исследований М. Камшиловым, в особенности над мутацией безглазия (eyeless) дрозофилы. Из результатов этих исследований в данной связи очень большое значение имеет экспериментальное доказательство возможности полного извращения реакции. Нормально мутация безглазия получает наиболее типичное выражение при культивировании на влажном субстрате. Путем искусственного отбора на максимальное выражение редукции глаза в культуре на подсохшем корме удалось добиться того, что мухи, выводившиеся на влажном корме, обладали большим числом фасеток глаза, чем мухи, выводившиеся на подсохшем субстрате. Если в исходной линии уменьшение влажности связано с увеличением размеров глаза (т. е. уменьшением выражения мутации), то в экспериментально полученной линии уменьшение влажности ведет к уменьшению числа фасеток (усиление выражения мутации). В результате искусственного отбора получена, следовательно, линия, обладающая иной зависимостью морфо-генетического процесса (развития глаза) от внешнего фактора (влажности субстрата, чем исходная форма. Интересно, что имеющаяся зависимость может быть не только изменена, но и уничтожена. При чередующемся отборе по тому же выражению безглазия на влажном и на подсохшем субстрате удалось добиться такого наследственного изменения линии, что эта мутация получала в обоих случаях одинаковое выражение. Признак, зависящий от внешнего фактора (влажности субстрата), превратился в независимый. Характерная форма реагирования мутации eyeless оказалась погашенной через искусственный отбор малых наследственных изменений.
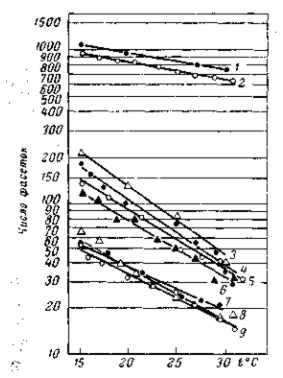
Рис. 25. Зависимость числа фасеток глаза нормальной дрозофилы и гомозиготных мутаций Ваг и Ultra-Bar от температуры. 1 — нормальная «дикая» форма; 2 — мутация, обратная к норме (получена в серии Ваг); 3—культура Ваг, не подвергавшаяся отбору; 4–6 — культуры Ваг, подвергавшиеся легкому отбору на уменьшение числа фасеток; 7–9 — разные линии Ultra-Bar. Температурная реакция этих мутаций имеет такой же характер, как и у нормальных мух, но выражена гораздо резче (норма более стабильна): если у нормы при повышении температуры от 15 до 30° число фасеток падает от 1000 до 800, то у Ваг оно при тех же условиях падает примерно от 200 до 40 (т. е. в 5 раз). По А. Г. Гершу, 1934
К вопросу о значении этого обстоятельства мы еще вернемся в следующем разделе (4). Здесь же отметим, что формы реагирования мутаций на изменения в факторах внешней среды могут быть в процессе отбора преобразованы. Следовательно, вредные выражения таких зависимостей могут быть либо погашены, либо видоизменены и в результате получат иное, быть может, благоприятное выражение.
Исследования М. Камшилова показали, кроме того, возможность установления зависимостей иного рода между развивающимся организмом и внешней средой. В опытах охлаждения четырехдневной личинки до 0° в продолжение 24 часов наблюдалась довольно значительная смертность. Повторение этого опыта в течение целого ряда поколений привело через естественный отбор к созданию линии дрозофилы, весьма устойчивой по отношению к такому охлаждению. Однако при этом был достигнут еще и следующий результат: в этой новой «холодоустойчивой» линии «приспособление» к охлаждению достигло такого уровня, что ее жизнеспособность при непродолжительном охлаждении личинки (в четырехдневном возрасте) оказывалась даже более высокой, чем без охлаждения.
Рис. 26. Температурная зависимость числа фасеток глаза у гомозиготных самок (А'А') и Infra-Bar (AA) и у соответствующей гетерозиготы (АА').Для графика применены логарифмы числа фасеток. Обе мутации дают прямо противоположную реакцию на изменение температуры. Однако у гетерозиготы не только не происходит взаимного погашения противоположных выражений этой зависимости, но характерная для Ваг обратная зависимость от температуры выражена даже еще резче, чем у гомозиготы. По А. Г. Гершу, 1930
Это показывает, что естественный отбор особей, развивающихся в определенных условиях внешней среды, приводит к установлению новых зависимостей между развивающимся организмом и факторами внешней среды. В некоторых случаях типичной смены факторов эта смена становится более или менее необходимым условием нормального развития организма. Вместе с тем и неблагоприятный фактор внешней среды может получить положительное значение в развитии организма (Камшилов, 1939а, б, 1941).
Все это подчеркивает возможность довольно быстрой перестройки зависимостей между развивающимся организмом и факторами внешней среды и показывает лишний раз, что реакции ор ганизма на изменения во внешних факторах имеют историческую базу. Благоприятные формы реагирования на внешние факторы не даны как изначальные свойства организма, а являются результатом естественного отбора особей в конкретных условиях развития организма в определенной внешней среде. Новые, элементарные формы реагирования любой новой мутации (как часть ее нормы реакций) обычно не имеют положительного значения. Однако в процессе естественного отбора (малых мутаций — модификаторов) эти формы реагирования могут менять свойхарактери быть либо погашенными, либо преобразованными в благоприятные, т. е. адаптивные, формы морфоге-нетических реакций.
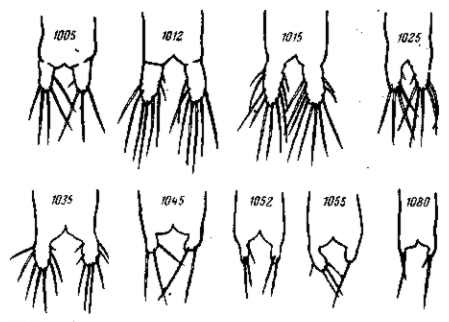
Рис. 27. Модификация Artemia salina в солевых растворах разной концентрации (на рисунке цифрами обозначены удельные веса). Культура проходит при уд. весе 1015 в явно оптимальных условиях — развитие фурк и щетинок достигает максимальной полноты. В наиболее концентрированных растворах фурки и щетинки совсем не развиваются. По A. Abonyi, 1915
б. Явление недоразвития в ненормальных условиях. На грани морфозов и адаптивных модификаций стоят явления недоразвития, нередко наблюдаемые в тех случаях, когда организм попадает в ненормальные условия внешней среды.
Мы только что упомянули, что в течение исторического развития организма (иногда даже в течение сравнительно кратковременного эксперимента) в определенной внешней среде, в особенности при наличии известной смены факторов, устанавливаются нередко зависимости такого рода, что некоторая последовательность в смене внешних факторов становится обязательным условием для нормального развития организма.
Такие зависимости весьма обычны для организмов с точно определенным, сезонным циклом размножения. Яйца многих насекомых, а также мелких ракообразных требуют зимнего охлаждения для начала их развития. То же самое относится иногда и к другим стадиям развития, —иначе развитие не возобновляется после «диапаузы». Особенно это характерно для растений, требующих для полного развития и созревания известного предварительного охлаждения для прорастания и весеннего пробуждения, а также определенных доз света, притом в виде светового дня соответствующей длительности. Отсутствие во внешней среде необходимых условий для индивидуального развития приводит такие организмы к явлениям недоразвития. Во многих случаях это означает отсутствие цветения у растения в чуждом климате или отсутствие половою созревания у животного. Организм не может противостоять таким изменениям и в новой среде не размножается. В других случаях явления недоразвития выражены не столь резко. Организм размножается, но оказывается соответственно модифицированным. Такие модификации нельзя назвать адаптациями, хотя они и сопутствуют переживанию организма в измененных условиях внешней среды.
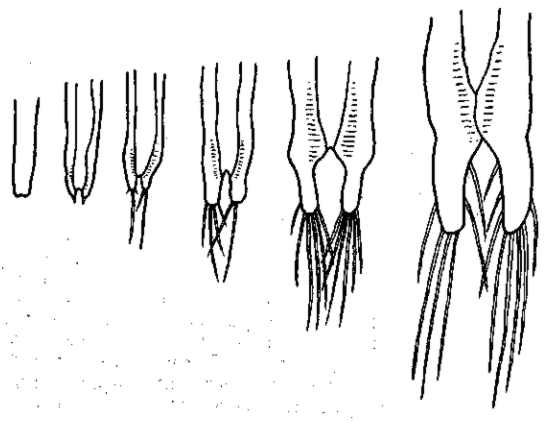
Рис. 28 Стадии индивидуального развития фурк и щетинок у Artemia salina в оз. Майли-Сор (Сев. Казахстан) при содержании С1 в воде около 6 : 100 и уд. весе около 1,10. По О. И. Шмальгаузен
Так, например, многие сухопутные растения, попадая в воду, не гибнут в ней, а продолжают расти. Они, однако, модифицируются, и такие изменения имеют характер общего недоразвития (lingerer). Соленоводный жаброног Artemia salina может жить и развиваться в очень соленой воде, приближающейся к насыщению. Однако он модифицируется в ней, изменяясь именно в сторону общего недоразвития — размеры рачка значительно уменьшаются, последние членики брюшка и фурка сохраняют личиночное строение (рис, 27, 28). Половое созревание можно считать преждевременным и всю эту ультрагалинную модификацию рассматривать как частный случай неотении (Abonyi, 1915). Морская мизида Mysis oculata, попадающая у нас на севере в условия опресненной воды, уменьшается в размерах и приближается к пресноводной Mysis relicta. Такое же измельчание наблюдается и при переходе морского таракана Chiridothea sibirica в устья рек и в пресные озера. Пресноводные модификации С. entemon-C. vetterensis отличаются уменьшенными размерами и узким тельзоном. Двустворчатый моллюск — мидия Mytilus edulis — значительно уменьшается в размерах, переходя в опресненные воды (например, Ботнического залива). То же самое касается многих других морских животных, которые при переходе в пресные воды приобретают карликовые размеры и недоразвитые формы.
Б. АДАПТИВНЫЕ МОДИФИКАЦИИ
В предыдущем разделе мы рассмотрели элементарные реакции организма, попадающего в совершенно новые условия внешних факторов, или реакции мутантного организма, которые также связаны с совершенно новыми взаимоотношениями между организмом и факторами внешней среды. Теперь мы познакомимся с уже установившимися в предшествующей истории формами реагирования. Некоторые из них имеют общефизиологический характер. В процессе эволюции они лишь вводятся в известные пределы оптимальных изменений. Таковы изменения скорости роста и общей величины тела, плодовитости, вегетации и цветения у растений в зависимости от питания, условий влажности и температуры. Другие реакции имеют более специальный (но в основе, конечно, также физиологический) характер, выражаясь в специфических приспособлениях организма и его органов к известным изменениям в физических факторах внешней среды. Мы их называем физиоген-ными модификациями. Например, изменения поверхности листьев в зависимости от интенсивности освещения, изменения пигментации у животных или адаптивные реакции в органах терморегуляции при изменении температуры. Наконец, третьи реакции выражаются в функциональном приспособлении органов и представляют, следовательно, лишь косвенный ответ организма на изменения в факторах внешней среды. Эта последняя категория функциональных модификаций может лишь условно быть отграничена от физиогенных модификаций, причем мы здесь отмечаем лишь относительно большую активность самого организма (рис. 29). АПТИВНЫХ РЕАКЦИЙ 291
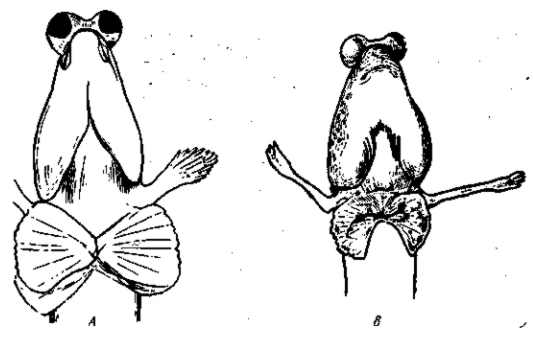
Рис. 29. Модификация прибрежной рыбы Periophthalmus variabilis А — нормальная водная форма; В— наземная модификация, полученная в эксперименте применением тироксина. По I. W. Harms, 1934
а. Модификации общефизиологического характера. Естественно, что возможности роста ограничиваются имеющимися запасами питательных материалов (у зародыша) и их дальнейшим поступлением при самостоятельном питании развивающегося организма. Точно так же понятно, что и верхний предел скорости роста ограничивается нередко возможной интенсивностью питания. Однако в процессе эволюции устанавливаются известные оптимальные нормы роста, определяемые экологическим положением организма и, в особенности, его соотношениями с другими организмами. Модификации возможны тогда лишь в сравнительно узких пределах этой нормы. В качестве приспособления мы должны здесь рассматривать не самую модификацию, а ее ограничение известными пределами. То же самое касается и скорости роста организма, которая имеет свой максимум. Минимум скорости роста находится в непосредственной зависимости от питания. Одаако сниженная скорость роста может быть компенсирована при позднейшем усилении питания. В этих явлениях компенсации мы также видим приспособление организма к восстановлению нормальных размеров при их нарушении вследствие временной недостачи питательных материалов. Плодовитость организма таюке зависит от питания. Однако уровень плодовитости определяется и экологическими условиями, т. е. положением организма в условиях его борьбы за существование. Адаптивной является и в этом случае не сама плодовитость и ее модификации, а именно ее ограничение известным оптимальным для данного вида уровнем.
Общая поверхность листьев растения и развитие его корневой системы подвержены также значительным модификациям, определяемым освещением и влажностью. В основе этих модификаций лежат также явления роста, определяемые условиями питания (ассимиляции). Адаптивный характер имеет опять-таки ограничение роста оптимальными в данных условиях пределами.
Во всех этих случаях мы имеем дело с общефизиологическими процессами, которые оказываются всегда очень изменчивыми вследствие постоянного обилия малых «физиологических» мутаций, выражающихся в изменении интенсивности обмена веществ, скорости роста, его длительности, а следовательно, и в изменении нормальных его пределов. То же самое касается и плодовитости, ее зависимости от питания и ее нормального и предельного выражения. Все эти зависимости явно меняются и в отдельных мутациях, и в процессе эволюции, определяемом естественным отбором оптимальных форм реагирования и пределов роста, оптимальной плодовитости и ее возможных пределов при разных условиях питания (температуры, влажности, освещения и т. п.).
б. Физиогенные модификации. Яснее выражается адаптивный характер модификаций, связанных более непосредственно с изменениями в физических факторах внешней среды. Так, например, в зависимости от условий освещения и влажности меняется величина, а нередко и форма листовой пластинки растений. Развиваются световые и теневые формы листьев, которые обладают оптимальной структурой для ассимиляции на прямом солнечном свету или в тени и вместе с тем соответствуют различным условиям транспирации. Необходимость ее ограничения при недостатке влажности ведет к дальнейшим адаптивным преобразованиям световых листьев в ксерофитные. Световые листья не только мельче и, как правило, менее рассечены, но обладают и более толстой кутикулой и узкими межклетниками, а также частыми, но более мелкими устьицами. Ксерофитные листья обладают и другими приспособлениями для удержания воды в тканях и для защиты от чрезмерной инсоляции (например, опушение листьев). Все эти приспособления весьма изменчивы. У некоторых ксерофитов прилистники или часть листьев преобразуются в колючки, но при развитии в условиях достаточной влажности вместо колючек вновь развиваются листья.
Во всех этих случаях модификации осуществляются через зависимость процессов роста и формообразования от условий освещения и степени влажности. У растений процессы роста сильно; зависят от вакуолизации растущих клеток: поэтому обеспеченность водой и ограничение испарения естественно способствуют более свободному росту растения в тени. С другой стороны, недостаток влаги в наземных частях и обильное испарение на солнечном свету ограничивают этот рост. Таким образом, адаптивные модификации здесь осуществляются на базе таких же общефизиологических процессов, как уже рассмотренные. В данных случаях они имеют лишь более локализованный характер и дополняются зависимыми же формообразовательными реакциями, основанными в значительной мере на неравномерном росте частей и на изменениях в гистологической дифференцировке. Формообразование; у растений вообще весьма зависимо от физических факторов,-а мутации, выражающиеся в изменении формы листьев и степени их расчлененности, принадлежат к наиболее обычным мутациям растительных организмов. Фенотипическая изменчивость таких мутаций и лежит, очевидно, в основе исторического развития способности к указанным адаптивным модификациям. У животных некоторым аналогом таких зависимостей могут служить температурные реакции млекопитающих, выражающиеся в изменении величины поверхности тела, его придатков и в изменении шерстного покрова. Эти реакции основываются также на широко распространенных физиологических процессах, связанных с общими закономерностями роста. Высокая температура способствует ускорению клеточных делений, а следовательно, повышению скорости роста при одновременном, однако, уменьшении размеров клеток, ускорении гистологической их дифференцировки, сокра^ щении дефинитивных размеров тела и увеличении его относительной поверхности. Об этом свидетельствует целый ряд экспериментальных данных по личинкам амфибий, эмбрионам птиц.
Мутации и расовые различия, сопровождающиеся изменением температурных реакций (пигментообразование) в волосах горностаевого кролика, по Ильину (см. рис. 8), вообще говоря, известны так же, как известны и наследственные изменения размеров тела,, размеров конечностей и придатков и, в особенности, длины, толщины, числа волос и формы их роста. Однако мутационные изменения зависимости роста этих частей от температуры, по-видимому, не изучены.
Температурные реакции животных выражаются гораздо чаще в изменении пигментации, однако такие модификации имеют лишь редко явно адаптивный характер. В особенности у птиц известны закономерные изменения окраски (правило Глогера, по которому в теплых и влажных местностях интенсивность пигментации возрастает, а в холодных и сухих окраска ослабляется,— это правило подтверждалось Алленом, Гёрницем, П. Серебровским и Реншем). Отчасти, по крайней мере, такие изменения могут быть модификационными, как это показывают некоторые эксперименты. Адаптивность этих изменений, однако, сомнительна. Пигментообразование — сравнительно простой физиологический процесс, зависящий от течения ферментативных окислительных реакций. При их ускорении под влиянием повышенной температуры окислительные процессы идут дальше и ведут к образованию более темных пигментов (черных эвмеланинов и желто-красных феомеланинов). В таком случае некоторое побледнение окраски у животных холодных стран, а также у пустынных животных может быть отнесено частично и за счет модификаций, адаптивность которых является лишь побочным результатом физиологических изменений процесса пигментообразования.
Однако известны и такие изменения окраски, которые имеют явно приспособительный характер, согласованный с изменениями общего фона и освещения окружающей среды. Это касается куколок некоторых бабочек, некоторых рыб, амфибий и рептилий. В большинстве случаев масштаб этих изменений весьма незначителен. Однако у некоторых позвоночных способность изменять свою окраску так развита, что издавна обращала на себя внимание натуралистов. Она основывается главным образом на сокращении отростков и на перемещении хроматофоров, реже — на перемещении пигментных зерен в отростках хроматофоров и иногда — на усиленном их размножении. Только последнее, имеющее место у молодых саламандр, может рассматриваться как зависимый морфогенетический процесс. Характер зависимости, однако, не установлен. В других случаях изменение окраски оказывается легко обратимым процессом перемещения или сокращения хроматофоров, осуществляемым через нервную систему в результате зрительных восприятий животного. Сюда относятся известные защитные реакции камбалы, окрашивающейся под цвет песка или гравия, на котором она располагается, а также классический пример хамелеона, меняющего свою окраску с поразительной скоростью и достигающего очень значительного разнообразия цветов (от светло-серого до темно-зеленого). Ясно, что эти приспособительные изменения относятся к группе легко и быстро обратимых физиологических реакций, которые мы оставляем здесь без рассмотрения (так же как и всю большую область явлений приспособительных двигательных реакций организмов — рефлексов, инстинктов и поведения животных).
К категории физиогенных модификаций физиологического характера следует отнести и изменение количества эритроцитов, а также содержания гемоглобина в крови позвоночных животных, в зависимости от атмосферного давления. Заметное увеличений этих показателей установлено у ящериц и млекопитающих, в частности и у человека, при их перемещении в горные местности (или при экспериментах в барокамере). Эти изменения бесспорно адаптивны. Они устанавливаются довольно быстро и полностью обратимы. По-видимому, они определяются парциальным давлением кислорода в самой крови и соответственно представляют одно из широко распространенных явлений функциональных компенсаций, которые часто проявляются при недостаточности некоторых жизненных функций.
в. Функциональные модификации. Разделение модификаций на общефизиологические, физиогенные и функциональные имеет лишь весьма условный характер, так как все они определяются физиологическими процессами, т. е. основываются на жизненных функциях организма. Поэтому можно было бы и все модификации, так же как и все адаптивные физиологические реакции, обозначить как функциональные приспособления в широком смысле, или аккомодации. Для наших целей, однако, удобнее понятие аккомодации дифференцировать. В данном случае мы обращаем внимание на различную активность организма. Функциональными в узком смысле слова мы будем считать адаптации, возникающие в связи с изменениями активности самого организма.
Если физиогенные адаптации характерны главным образом для растений и осуществляются через посредство возникающих, еще не дифференцированных тканей, то функциональные адаптации свойственны в основном животным и осуществляются путем перестройки дифференцированных уже и нормально функционирующих тканей и органов. Они определяются всегда ролью известной ткани или органа в жизнедеятельности организма в целом (рис. 29).
Во всех таких адаптивных реакциях усиленная функциональная нагрузка ткани или органа ведет к гипертрофии, а систематическая недогрузка — к ослаблению органа и явлениям атрофии. К сожалению, физиологический механизм этих изменений не изучен в достаточной мере. Если нормальное функционирование органа связано с поддержанием известного равновесия между процессами диссимиляции и ассимиляции, то в состоянии покоя или при перенапряжении органа происходит, по-видимому, не столько нарушение количественной стороны этого равновесия, сколько изменение его качественной характеристики. При отсутствии специфических продуктов разрушения данного органа нет и восстановления его специфических элементов. Обмен веществ ограничивается одним лишь основным обменом. Обмен функционирующего органа имеет, очевидно, не только количественно, но и качественно иной характер. Специфические продукты обмена функционирующего органа являются, вероятно, стимуляторами процессов физиологической регенерации дифференцированных элементов органа. Увеличение концентрации этих продуктов выше известного уровня ведет к преобладанию восстановительных процессов над функциональным распадом. Функциональная гипертрофия приводит к установлению нового равновесия па более высоком уровне концентрации специфических продуктов обмена. Наиболее характерны явления функциональной гипертрофии в мышцах. Тренировка мышц ведет к их усилению вследствие увеличения толщины отдельных мускульных волокон. Сила мышцы, как известно, пропорциональна ее поперечному сечению. Сходны и явления функциональной гипертрофии в нервной ткани. И здесь они связаны с увеличением массы нервных клеток и увеличением поперечника их нервных отростков. Однако у зародыша или личинки ЕОЗМОЖНО и увеличение числа нервных клеток и, следовательно, и нервных волокон. Это видно из ряда экспериментальных исследований (над амфибиями).
Рис. 30. Структура верхней части бедреннод кости человека. Костные пластинки располагаются в губчатом веществе кости по линия:» максимального натяжения и сжатия (Wolff, 1892)
В опорных тканях реакция на увеличение механической нагрузки выражается и в увеличении объема, и в перестройке самой структуры, если эта нагрузка меняет свое направление. Эти реакции наиболее изучены на волокнистой и эластической соединительных тканях и на костной ткани. Адаптивность изменений строения опорных тканей исключительно высока. В соединительных тканях она выражается в развитии обыкновенных и эластических волокон по линиям натяжения. В костной ткани она выражается в развитии более сложной внутренней структуры в виде системы костных пластинок, располагающихся главным образом по линиям сжатия и линиям растяжения. Там, где эти направления расходятся, образуются более сложные губчатые структуры с перекрещивающимися костными перекладинами (рис. 30).
Исключительно пластична кровеносная система. Она в высшей степени быстро перестраивается согласно функциональным требованиям возросшей массы тканей или их возросшей активности. Даже во взрослом организме легко устанавливается коллатеральное кровообращение при облитерации отдельных сосудов.
Такая же легкая приспособляемость обнаруживается и во многих внутренних органах. Известна высокая способность сердечной мышцы к тренировке РГ К компенсации при нарушениях нормального кровообращения. Точно так же возможны и адаптивные изменения дыхательной поверхности жабер у личинок земноводных (аксолотля) и дыхательной поверхности легких у наземных позвоночных. Весьма пластичны и почки позвоночных. Даже у млекопитающих возможны новообразование вторичных почечных канальцев у взрослого животного и далеко идущие явления компенсации при усиленной функциональной нагрузке, падающей на одну или обе почки.
Во всех этих функциональных адаптациях или компенсациях имеется нечто общее. Самое функционирование связано с процессами, ведущими к поддержанию данного органа, ткани или системы на известном уровне развития, и оно же ведет к превышению этого уровня при повышенной нагрузке. По всей вероятности, такого рода функциональные зависимости устанавливаются вместе с возникновением данной дифференцировки в процессах взаимной обусловленности исторического развития органа (ткани, системы) и его функции. Это историческое развитие всегда поддерживалось, с самого начала данной дифференцировки, взаимной обусловленностью индивидуального развития структуры органа (системы, ткани) и его функции. Можно думать, что и эволюция таких структур шла в значительной мере на базе этой взаимообусловленности структур и функций в индивидуальном формообразовании (см. дальше раздел Г. Значение адаптивных модификаций в эволюции. Стр. 305).
Иными словами, мы полагаем, что новые функциональные дифференцировки возникают всегда на базе жизнедеятельности самого организма. В связи с новым расчленением функций устанавливаются и новые структуры как результат функциональной деятельности организма. Функциональные дифференцировки возникают под влиянием самой функции, а в дальнейшей эволюции происходит их стабилизация и включение через смену морфогенных факторов в число автономно развивающихся структур. При таком понимании эволюции активных функциональных структур вполне понятно и не требует специальных пояснений то обстоятельство, что эти структуры фиксируются сразу же вместе с теми физиологическими механизмами (специфика обмена веществ), которые сопровождают их возникновение. Сопутствующие условия развития становятся в процессе эволюции необходимыми условиями реализации известных структур (см. стр. 54 о внешних условиях развития). Продукты специфического обмена получают при этом значение морфогенных веществ, стимулирующих развитие именно данных структур.
К функциональным адаптациям можно отнести также образование мозолей, т. е. утолщений рогового слоя кожи, наблюдаемых у птиц и особенно у млекопитающих, в тех частях общих покровов, которые испытывают постоянное давление или трение. Такие реакции можно рассматривать также как явления функциональной гипертрофии, основанной на стимуляции процессов физиологической регенерации.
Функциональным адаптациям мы приписываем очень большое значение в возникновении новых дифференцировок в процессе эволюции высших позвоночных. Как это осуществляется, мы рассмотрим несколько далее. Однако отметим тут же, что это понимание генезиса функциональных дифференцировок подводит нас в то же время и несколько ближе к пониманию самого возникновения способности к функциональным модификациям.
г. Индивидуальная коадаптация органов. Индивидуальная приспособляемость организма выражается в физиологических и морфологических изменениях, связанных с изменениями в факторах внешней среды. Собственно физиологических реакций мы здесь не касаемся, как бы велики ни были их адаптивности и их значение в эволюции. Что же касается морфологических адаптации, выражающихся в более стойких изменениях формы и структуры организма и его органов, они, как сказано, могут иметь зависящую от внешних факторов общефизиологическую основу (явления роста и размножения). Они также могут строиться на частных зависимостях формообразовательных процессов от внешних факторов (температуры, света, влажности) или основываться на их зависимости от функциональной нагрузки (диктуемой, очевидно, положением организма во внешней среде).
Части лабильного организма и его органы приспособляются, однако, не только к внешней среде. Можно говорить и о взаимном приспособлении органов внутри организма. И в этом случае речь может идти о взаимозависимостях в общефизиологических процессах роста, ведущих к изменениям в пространственных соотношениях, о взаимозависимостях в специфических факторах морфогенеза (морфогенетические корреляции) или о взаимозависимостях в функциях (эргонтические корреляции). Все эти связи могут вести к согласованным преобразованиям в соотношениях органов и их частей (если они имеют регуляторный характер).
Наличие таких взаимозависимостей различного рода представляет эмпирический факт. Мы различаем геномные, морфогенети-ческие и эргонтические корреляции (Шмальгаузен, 1938а, 1939а, 1942). Из них часть морфогенетических и, по-видимому, все эргон-тические корреляции имеют регуляторный характер. Это означает, что изменение одного звена (зачатка, ткани, органа) влечет за собой согласованное изменение другого звена (зачатка, ткани, органа) корреляционной цепи. Морфогенетические и эргонтические корреляции регуляторного характера строятся на базе элементарных морфогенетических взаимозависимостей, проявляющихся уже в отдельных мутациях (выражения множественного и многостепенного плейотропизма см. далее гл. 4). На отдельных мутациях и на морфозах нередко уже ясно видны взаимозависимости в изменении соотношения органов. Они изучались в последнее время Рапопортом (1941) на хемоморфозах и М. Камшиловым, а также Р. Берг на мутациях с изменчивым выражением. М. Камшилов установил существование зависимостей между различными выражениями одной мутации. Путем искусственного отбора, а также при разных условиях развития достигался сдвиг в виде закономерного ряда последовательных изменений различных частей (органов) . М. Камшилов видит в этом указание на существование взаимозависимостей типа градиентов. Р. Берг (1943) пришла к такому же выводу. И. Рапопорт предпочитает говорить о морфогенетических взаимозависимостях в более общей форме.
Во всяком случае, эти исследования показывают, что в мутациях обнаруживается наличие корреляций и, соответственно, в них происходит взаимозависимая перестройка частей. Такая же взаимозависимость обнаруживается и в морфозах. В процессе эволюции такие элементарные взаимозависимости приобретают целесообразно-регуляторный характер. Как уже сказано, многие морфогенетические взаимозависимости (корреляции) в развитии частей нормального организма (как они изучаются «механикой развития») имеют именно регуляторный характер. Такой же характер имеют, по-видимому, и все эргонтические корреляции. Эргонтические корреляции основываются на тех же реакциях, что и функциональные адаптации. Механизм адаптации и коадап-тации органов здесь по существу совпадает. Разница лишь в источнике изменения — лежит ли он во внешней среде или в изменении другого органа. Если увеличение количества эритроцитов и гемоглобина, увеличение сердечной мышцы или увеличение дыхательной поверхности легких определяются уменьшением парциального давления кислорода в крови, то по сути совершенно безразлично, что является первоисточником этих изменений — пониженное атмосферное давление в горной местности или повышенная активность организма в связи с трудностью добычи необходимого питания или при переходе к иному способу движения, требующему более значительной работы мышц (например, при летании). Если увеличение числа почечных канальцев определяется повышенной концентрацией продуктов обмена в крови, то опять-таки механизм этого изменения один и тот же как в том случае, если это увеличение зависит от изменения качества пищи или от изменения активности организма и количества пищи, так и в том случае, если оно зависит от более глубокого изменения самого обмена или от простого увеличения массы самого животного.
Практически явления адаптации организма к внешней среде и явления коадаптации органов неразграничимы, так как любая адаптация известных частей к изменению в факторах среды неизбежно сопровождается и соответственной коадаптацией, т. е. взаимозависимым изменением других частей и, строго говоря, всей организации. Точно так же и наоборот: любая перестройка организации меняет положение организма во внешней среде и изменяет приспособления отдельных частей к различным условиям этой среды.
Наличие системы корреляций регуляторного характера (об ее происхождении см. главу 4 этого раздела) приводит к тому, что изменение в процессе эволюции (или даже в результате индивидуальной адаптации) одной части или органа влечет за собой согласованное изменение другой части или органа. Такое согласование происходит отчасти именно в процессе индивидуального развития. Взаимное приспособление органов достигается путем согласования пространственных соотношений через морфогене-тические взаимозависимости, коррелятивный рост и функциональные взаимозависимости. Благодаря этому приспособления организма к окружающей среде приобретают целостный характер. Индивидуальная адаптация известных органов к измененным факторам внешней среды дополняется такой же коадаптацией других органов, так что в результате происходит согласованная перестройка всей организации. Можно говорить об установлении новой адаптивной нормы.
Однако система морфогенетических корреляций регуляторного характера достигает большой сложности только у позвоночных животных. Поэтому адаптации растений и большинства животных далеки от такой совершенной внутренней согласованности — они ограничиваются нередко частными изменениями отдельных органов. Впрочем, для организмов, живущих на грани различных сред и попадающих в весьма различные условия существования, даже физиогенные адаптации могут (главным образом через систему геномных корреляций) приобрести целостный характер вполне выработанных «адаптивных норм».
В. АДАПТИВНЫЕ НОРМЫ И ИХ СМЕНА
В организмах со слабым развитием системы регуляторных корреляций коадаптация частей осуществляется лишь медленно, в порядке естественного отбора наиболее жизненных, наиболее гармонических соотношений.
Изменение отдельной части или признака организма соответственно изменению фактора внешней среды нарушает до известной степени гармоническую согласованность частей. Этим самым значительно снижается жизненная и селекционная ценность любого положительного изменения форм реагирования. Поэтому способность к частным адаптациям дополняется по меньшей мере и другими реакциями, т. е. некоторыми зависимостями, позволяющими внести большую согласованность в изменения всего организма в целом. Такие зависимости имеют на первых ступенях своего развития характер геномных корреляций. В процессе эволюции вырабатываются, таким образом, типичные реакции, ставящие весь организм в новое положение. Между типичными формами приспособления образуются разрывы, и переход от одной типичной формы к другой приобретает характер скачка. В особенности это имеет место при резком изменении положения во внешней среде и доминирующем значении физиогенных адаптации. Поэтому такие переходы от одного типа приспособления к другому более ярко выражены у растений, живущих на грани различных биотопов и попадающих систематически и случайно то в один, то в другой биотоп. При развитии типических норм реагирования организма, в виде различных типов приспособления, особое значение приобретают регуляторные процессы.
При небольшой интенсивности внешнего фактора может не наступать никакой видимой реакции. На известном уровне этой интенсивности, соответствующем нижнему порогу реактивности организма, наступает сразу в полном своем выражении реакция, зависимая от данного фактора. Дальнейшее изменение интенсивности не изменяет результата формообразовательных процессов, вплоть до некоторого уровня, соответствующего верхнему порогу реактивности, по достижении которого данная реакция вновь выпадает, заменяется морфозом или совершенно иной адаптивной реакцией. В особенности интересны, конечно, случаи полного переключения одной реакции на совершенно иную, но также приспособительную реакцию.
Между нижним и верхним порогами нормальной реактивности организма, выражающейся в реализации адаптивной модификации, лежит широкий диапазон возможных изменений интенсивности, на которые организм как будто вовсе не реагирует. Это указывает на частичную утрату детерминирующего значения внешнего фактора. Последний выступает в роли стимулятора, лишь возбуждающего известную цепь формообразовательных процессов, которые протекают в основном под влиянием внутренних факторов развития. Повышение интенсивности внешнего фактора за пределы нижнего порога реактивности организма не оказывает заметного влияния на течение этих процессов вследствие очевидного наличия регуляторных механизмов, противодействующих этим влияниям. Такую форму индивидуального развития, зависящего в своем осуществлении от известного минимума интенсивности внешнего фактора, который дает лишь первый толчок к развертыванию внутреннего механизма типично детерминированного формообразовательного процесса, мы называем авторегуляторным развитием.
Процессы авторегуляции представляют адаптивные ограничения нормальной реактивности известными пределами. С таким ограничением реактивности известными пределами «нормы» мы познакомились уже при рассмотрении общефизиологических реакций (в явлениях роста). Возникновение подобных регуляторных механизмов на базе элементарных реакций, свойственных отдельным мутациям, нетрудно себе представить, так как такие реакции практически всегда укладываются в известные рамки, которые могут сдвигаться как в одну, так и в другую сторону при комбинировании разных мутаций и в процессе естественного отбора различных комбинаций.
Типичная окраска горностаевого кролика (рис. 8) — белый с черными носом, ушами, хвостиком и концами лапок — развивается, по Н. Ильину, нормально при температуре среды от 2 до 14°. При температуре выше 14–16° концы лапок становятся белыми. В пределах от 2 до 14° окраска не зависит от изменений температуры среды. Необходим минимум в 2° для того, чтобы типичная окраска горностаевого кролика проявилась в полной мере; 2° являются нижним пороговым уровнем нормальной окраски, а 14°— верхним порогом нормальной реактивности горностаевого кролика на температурные изменения. При более высокой температуре, как сказано, развиваются белые лапки. В пределах от 16 до 25° мы имеем также вполне типичную и устойчивую вторую «норму» —- белый кролик с черными ушами, носом и хвостиком. При температуре выше 25° начинается побеление ушей, а при 29° начинают белеть нос и хвостик. Такая высота различных порогов реактивности кожи горностаевого кролика характерна, однако, только для московской расы. У западных рас температурные пороги находятся на гораздо более высоком уровне (для кожи бока нижний порог белой окраски у московской расы + 2°, у польской + 11°, а у саксонской +21°). Имеются и индивидуальные наследственные различия пороговых уровней у отдельных кроликов.
Все это показывает возможность смещения пороговых уровней реактивности как в индивидуальных уклонениях (мутациях) и их комбинациях, так и в результате естественного отбора, ведущего к образованию различных наследственных форм.
Таким образом, подобные реакции вводятся в ограниченное русло более определенной «нормы», приспособленной именно к известным условиям внешней среды (например, к известным пределам температуры). При этом для разных условий могут вырабатываться и характерные для них и одинаково приспособленные различные «нормы» (для низких зимних температур одна норма окраски, например белая; для более высоких, летних температур — другая норма, например серая или бурая).
Известны многочисленные случаи сезонного полиморфизма с реакциями именно такого рода. Если куколка Araschnia (Vanessa) levana зимует или в эксперименте подвергается любым температурам ниже 0°, то развивается типичная рыжая весенняя форма А. levana. При температурах, превышающих + 2°, развивается типичная темная, летняя форма A. prorsa.
У животных адаптивный полиморфизм имеет именно почти всегда сезонный характер. В особенности характерны явления «цикломорфоза» у планктонных организмов — коловраток и дафний. Здесь формообразование зависит в своей специфике от взаимосвязанного комплекса изменений температуры и питания и выражает приспособление к парению в воде различного удельного веса (увеличение поверхности у летних форм).
Можно привести многочисленные примеры модификационного полиморфизма, связанного с существованием регуляторных механизмов, определяющих развитие нескольких приспособительных норм у растений. Как раз у растений, лишенных возможности активно перемещаться и переходить из одного биотопа в иной, более подходящий, большое значение имеет способность к физиогенным адаптивным модификациям и, в частности, нередко имеются хорошо дифференцированные целостные приспособительные нормы. Как уже упомянуто, это в особенности характерно для растений, живущих на грани резко различающихся биотопов. В этом случае регуляторный характер реакции, полностью реализующейся в типичном виде на известном уровне интенсивности внешнего фактора, выступает вполне ясно.
У водяной гречихи (Polygonum amphibium) при известной степени влажности (даже без непосредственного воздействия воды) внутренний механизм развития воздушных листьев сразу переключается на механизм развития водных «плавающих» листьев (Goebel, 1928). У стрелолиста (Sagittaria sagittifolia) при известном ослаблении освещения реакция образования наземных стоячих листьев переключается на развитие водных лентовидных листьев (рис. 31). Более сложный комплекс факторов действует при модификациях растений в горных местностях. Здесь при комбинированном влиянии интенсивной инсоляции и низкой температуры развивается характерный альпийский габитус (низкий стебель с прикорневой системой обычно опушенных листьев, глубокие корни). В менее яркой форме такая множественность реакций свойственна очень многим растениям, встречающимся на различных почвах и вообще в различных экологических условиях («эко-фены» Турессона).

Рис. 31. Изменение формы листьев на одном и том же экземпляре стрелолиста Sagittaria sagittifolia в зависимости от условий среды. Пример нескольких типичных адаптивных модификаций (норм). По Курсанову
Во всех этих случаях мы имеем явления модификационного полиморфизма, связанного с авторегуляторным типом развития. Каждая форма вполне приспособлена к известному комплексу внешних условий, реально встречавшихся и встречающихся в жизни данного вида в качестве случайных, периодических (сезонных) или локальных явлений. Вид в целом приспособлен не в одной своей форме, а в двух или нескольких формах, и не к одному сезону или биотопу с характерным для него комплексом условий, а к нескольким. Поэтому, в случае изменения среды, связанного с выпадением известного комплекса условий, характеризующего один из сезонов или один из биотопов, к которым вид был приспособлен, вид не вымирает, так как остается еще иной комплекс факторов, к которому данный вид был уже приспособлен. Заболачивание почвы, распространение леса или, наоборот, отступаний леса и развитие степи приводят к обязательной реализации тех реакций, которые раньше осуществлялись лишь случайно, периодически или локально (водная форма, теневая, ксерофитная и т. п.). Происходит «смена адаптивных норм», при которой прежняя главная реакция теряет свое значение и уступает место новой, бывшей раньше лишь второстепенной. Все реакции приобрели уже заранее целостный характер, и это обеспечивает немедленную перестройку всего организма даже при быстрых изменениях физических факторов внешней среды.
Прибрежное или болотное растение (рис. 32), иногда заливаемое водой и приобретшее уже способность развивать водные , листья, может в любое время перейти к чисто водной жизни. Растение континентального климата, теряющее иногда при засухе свои листья и приобретшее способность в этих случаях ассимилировать при помощи поверхностных тканей стеблей или листовых черешков, может при развившейся сухости климата быстро превратиться в типичного ксерофита, ассимилирующего при помощи стеблей и черешков (филлокладиев и филлодиев).
Эти представления о смене адаптивных норм можно было бы развить и дальше. Можно говорить о выделении новых норм, об увеличении числа норм, о расширении зоны адаптивности, о замене главной нормы второстепенной, о сокращении числа норм и сужении зоны адаптивности реакций (специализация). Однако значение самого принципа здесь освещено в достаточной мере, а детализация завела бы нас далеко за пределы разбираемой проблемы «факторов» эволюции.
Г. ЗНАЧЕНИЕ АДАПТИВНЫХ МОДИФИКАЦИЙ В ЭВОЛЮЦИИ
Вопрос о значении адаптивных модификаций в эволюции был в свое время правильно оценен Ч. Дарвином, хотя позднейшие «дарвинисты» видели в его высказываниях по этому поводу лишь ненужные «уступки» ламаркизму. Только у Л. Моргана, Болдуина и ряда других дарвинистов мы находим превосходную, до сих пор непревзойденную разработку этого вопроса с дарвинистических (в общем) позиций. К сожалению, позднейшие успехи генетики привели к распространению неодарвинистических представлений. Адаптивным модификациям при этом не отводилось вообще какого-либо места среди факторов эволюции. Только в новейшее время этот вопрос был вновь поставлен рядом советских авторов (Кирпичников, 1935, 1940; Лукин, 1935, 1936, 1939; Шмальгаузен, 1938а, 1939а, 1940а, 1941), которые рассматривали его с различных точек зрения. Однако и до сих пор вопрос о значении адаптивных модификаций не получил всестороннего освещения и, главное, не получил еще достаточной оценки. Я попытаюсь отметить здесь хотя бы основные положения для будущей его разработки. На первом месте я бы поставил значение самой лабильности, т. е. способности к адаптивным модификациям (более широкого вопроса об индивидуальной приспособляемости в физиологических реакциях и нервной деятельности я здесь не касаюсь) для жизни вида и для его потенциальной способности к эволюции. Затем я хочу отметить ведущее значение конкретных модификаций при изменении факторов внешней среды и при активном переходе в иную среду. Наконец, я не могу не обратить еще раз (Шмальгаузен, 1939а) внимания на значение способности к адаптивным функциональным модификациям в процессе установления действительно совершенно новых дифференцировок.
Рис. 32. Вполне развитые формы (адаптивные нормы) стрелолиста. По Schenck, 1915. а — наземная, б — водная форма
а. Модификационная эврибионтностъ. Способность к адаптивным модификациям означает всегда расширение среды обитания для данного вида организмов (или распространение на более значительный период времени в году — при сезонном полиморфизме). Организм может при соответствующих модификациях существовать и размножаться не в одном биотопе, а в двух или нескольких. Это означает но меньшей мере увеличение численности особей данного вида. Нередко это ведет и к более широкому распространению (в пределах возможной аккомодации). Вместе с тем это означает и уменьшение интенсивности элиминации, так как способность к индивидуальной адаптации предполагает и возможность приспособления к временным изменениям внешних факторов в пределах каждого из биотопов. Наконец, возможность известной адаптации и коадаптации означает до некоторой степени и сглаживание того нарушения соотношений, которое вносится теми или иными мутациями. Это связано опять-таки со снижением интенсивности элиминации.
Таким образом, в случаях развитой способности к адаптивным модификациям мы имеем (при прочих равных условиях) наличие более многочисленной популяции данного вида, живущей в более разнообразных условиях (биотопах), более сплошными массами (частично отпадает экологическая разграниченность биотопов) и в условиях менее острой элиминации физическими факторами ^при функциональных модификациях это касается и биотических факторов). Одним словом, результатом этой способности будет более или менее ярко выраженная эврибионтность, которую в этом случае можно ооозначить как «модификационную» эвриоионтность в отличие от «физиологической» эвриоионтности, о которой речь будет в дальнейшем.
Рис. 33. Закладка обыкновенного гребня (А) у 8-дневного зародыша домашней курицы и розовидного ( В) у 8,5-дневного зародыша породы виандот одинаковой стадии развития. Поперечные разрезы на уровне слезно-носового канала. Закладка розовидного гребня захватывает большую площадь покровов и значительную массу мезенхимы. Одновременно развивается более широкая носовая перегородка (sn). Закладка носовых костей запаздывает; с — обонятельный мешок, I — слезно-носовой канал, m — закладка гребня, п — закладка носовых костей
Значение морфологической приспособляемости организма не исчерпывается, однако, достижением более широких возможностей существования и размножения. Мы знаем, что в больших популяциях идет более легкое распространение и накопление мутаций и более свободное их комбинирование (панмиксия). Малая интенсивность прямой элиминации индивидуальных уклонений способствует также свободному накоплению многих мутаций и большему разнообразию возможных комбинаций.
В разнообразных условиях существования поддерживается всегда и известный гетероморфизм, т. е. генетическое многообразие форм.
Таким образом, лабильность организации, т. е. способность организма к приспособительным модификациям, естественно связывается всегда и с эволюционной пластичностью, т. е. мобильностью вида в процессе исторического преобразования наследственных форм. В этой повышенной пластичности, т. е. потенциальной способности к прогрессивной эволюции, мы и видим наиболее общее выражение значения индивидуальной приспособляемости организмов.
6. Смена климатических зон и биотопов. Способность к адаптивным модификациям означает некоторую потенциальную способность к миграции за пределы первоначального местообитания. Климатические факторы не отличаются постоянством, и если организм может приспособляться к временным их изменениям, то он способен мигрировать в такие климатические зоны, в которых эти случайные уклонения в факторах среды приобретают постоянное значение. Так, например, организмы, живущие в местности, поражаемой иногда засухой, и приспособляющиеся к этим условиям, могут перейти в более засушливую область, где такие засухи представляют нормальное явление. Организмы резко континентального климата, приспособленные к жизни в весьма различной температуре (эвритермные), могут легко перейти к жизни в более холодном или более теплом поясе страны с менее континентальным климатом. Точно так же возможна в известных пределах и миграция из одного биотопа в другой. В особенности легко осуществляется, конечно, переход из общей среды в частную. Если организм индивидуально приспособляется к жизни в нескольких оиотопах, то он легко может осесть в любом из этих биотопов. Конечно, это означает специализацию, т. е. превращение эврибиоита в стенобионта. Такой процесс идет с большой легкостью.
При исторических сменах климатов, с их экологическими и биоценотическими последствиями, индивидуальная приспособляемость приобретает исключительное значение, организм, обладающий способностью модифицироваться соответственно локальным или сезонным изменениям в факторах внешней среды, оказывается вполне приспособленным, если подобные изменения становятся постоянной осооенностыо среды. Условно можно было бы говорить о преадаптации, если бы этот термин не связывался с представлениями известной теории Кено. Здесь по сути имеет место, конечно, не предварительное приспосооление, а всегда лишь приспосооление к реальным условиям существования данного времени. Однако условия, которые в данную эпоху или в данной области встречаются лишь локально, время от времени или периодически, могут в следующую эпоху распространиться на новые области или приобрести постоянное значение. Тогда прежняя второстепенная адаптивная норма может приобрести значение главной или даже единственной нормы. Такое «приспособление» организма происходит с огромной скоростью — в течение развития одного поколения. Вслед за этим произойдет, конечно, и наследственная перестройка организма соответственно его новому положению во внешней среде. Это осуществляется исподволь в процессе стабилизирующего отбора. В данном случае адаптивная модификация получает ведущее значение. Именно конкретная адаптивная модификация намечает путь дальнейшей эволюции. Во всех этих случаях «приспособления» не возникает, однако, ничего нового. Организм отвечает на изменения во внешней среде определенной реакцией, способность к которой уже давно приобретена в течение предшествующей эволюции данного организма в изменчивой внешней среде.
В этой связи мы приписываем особое значение функциональным модификациям, так как они отличаются большой гибкостью и, кроме того, определяются не столько изменением физических факторов внешней среды, сколько изменением биотической обстановки. Наконец, в этом случае приобретает особую роль активность самого организма. При изменении внешней обстановки, а также в процессе эволюции организма, он активно меняет свое положение во внешней среде. Млекопитающие, переходящие в поисках пищи из леса в степь, встречаются с иными врагами и не имеют иных средств защиты от больших хищников, кроме быстрого бега (крупные животные) или зарывания в землю (мелкие). Это означает для конечностей смену функций, а для всего огранизма — сложный комплекс функциональных модификаций. Млекопитающее, живущее в лесу, может находить защиту на деревьях. Это связано с приспособлением конечностей для лазанья. Широкая способность к функциональным модификациям во всяком случае значительно облегчает такой переход к древесному образу жизни. Следовательно, на базе функциональных модификаций могут возникать и совершенно новые приспособления.
Способность к функциональным модификациям оценивается нами очень высоко. Мы полагаем, что на базе общих реакционных способностей могут выявляться все новые частные реакции, дающие действительно новое, не только количественно, но и качественно новое оформление приспособляющемуся организму. Таким образом, адаптивная модификация может выходить и за пределы общей приспособленности реакций, приобретенной в предшествующий период эволюции. Она может вести к новым частным дифференцировкам, которые получают такую же ведущую роль в дальнейшей наследственной перестройке организма.
в. Возникновение новых функциональных дифференцировок. Главным процессом прогрессивной эволюции организмов является дифференциация, основанная на разделении функций и состоящая в расчленении однородного целого на разнородные части. Дифференциация органически связана и с противоположным процессом интеграции (см. ниже, гл. 4, а также И. Шмальгаузен, 1938а, 1942). Она означает прогрессивное «освоение» факторов внешней среды. Организм отвечает все более дифференцированной реакцией на внешние факторы и, с другой стороны, по мере «освоения»!, воспринимает все более тонкие градаций этих факторов, устанавливая между ними и качественные различия. Он дифференцируется, и по мере этой дифференцировки увеличивается качественное многообразие воспринимаемых факторов внешней среды. Взаимоотношения между организмом и средой все более усложняются. Определяющим моментом в прогрессивной эволюции организма являются устанавливающиеся конкретные взаимоотношения между организмом и средой, выражающиеся в борьбе за существование и ведущие через естественный отбор к преобразованию организма. В эволюции низших форм жизни ведущим процессом является, очевидно, освоение все новых сторон внешней среды и вместе с тем изменение и усложнение среды, определяемое в свою очередь размножением и увеличением многообразия органических форм, усложнением связей и взаимоотношений, в конкретных биоценозах. По мере прогрессивной эволюции организма увеличивается, однако, его активность, и ведущая роль в изменении взаимоотношений с внешней средой переходит к самому организму, все более дифференцирующему свои реакции, охватывающему новые факторы среды, активно мигрирующему в новые биотопы, активно вырабатывающему новые средства защиты и активно переходящему, по мере необходимости, на новое питание. В этом случае значение индивидуальной приспособляемости (способности к аккоммодации) непрерывно возрастает. В частности, возрастает значение функциональных модификаций и в особенности возрастает значение физиологических аккомодаций и приспособительного поведения.
В функциональных модификациях возникают и новые соотношения частей, создаются новые нормы. Переход к иному образу жизни (или на деревьях и т. д.), к иному способу передвижения (бег, лазанье, плаванье и т. д.), к иной пище (питание насекомыми, водными животными, растениями, семенами и т. д.), к иным способам добычи пищи (подстерегание, хватание на бету или на лету челюстями или лапами и т. д.), к иным способам защиты (бегом, зарыванием, вооруженной защитой или пассивной и т. д.) связан с различной активностью тех или других органов и ведет отчасти непосредственно к функциональной перестройке организма (Шмальгаузен, 1942а, 1939а). Экспериментальные данные показывают возможность такой перестройки (рис. 29). Что же касается масштаба перестройки, достигаемой путем аккомодации, то это вопрос совершенно второстепенный. Важно то, что эта перестройка достигается очень быстро — в течение одного онтогенеза, т. е. гораздо быстрее любого изменения внешней среды или проникновения вида в новый биотоп. Уже небольшое приспособительное изменение ставит организм в благоприятствуемое положение, и если стабилизация новых соотношений произойдет с достаточной скоростью (а стабилизирующая форма отбора идет, очевидно, с гораздо большей скоростью, чем ведущая форма естественного отбора), то на этой базе возможна и дальнейшая адаптивная модификация, как следующая ступень приспособления вида к новым условиям существования. На основе смены функций возникают, таким образом, новые модификации, которые являются ведущими в дальнейшей, более медленной наследственной перестройке организации.
Однако мы должны здесь подчеркнуть и возможность возникновения совершенно новых дифференцировок в результате индивидуальной аккомодации организма. Все новые дифференцировки связаны с разделением функций и определяются различным положением частей по отношению к внешней среде или к другим частям того же организма. Именно на этой почве происходили все прогрессивные изменения организации. Конечно, всегда остается законным вопрос: насколько такие именения определяются непосредственно модифицирующим влиянием самой функции, насколько — отбором новых, более дифференцированных форм реакций на эту функцию и насколько — отбором «независимых» наследственных изменений данной структуры. Несомненно, это бывает различно в отдельных случаях. Мы отмечаем, однако, принципиальную возможность непосредственного влияния функции и возрастающее значение этого влияния по мере установления высших, более активных форм жизни, характеризующих эволюцию наземных позвоночных.
Активность высших животных определяется в значительной мере дифференциацией нервной и мышечной систем. Все остальное имеет подчиненное значение. Однако как раз в отношении нервной и мышечной систем роль тренировки исключительно высока. У птиц и, особенно, у млекопитающих тренировка нередко входит как обязательный элемент онтогенетического развития молодого животного (через игры и «обучение»). Мы знаем, что дифференциация функций между частями одной мышцы приводит всегда к их обособлению и, во всяком случае, комплекс однородных мышечных пучков может распасться на разнородные, вполне самостоятельные мышцы (такова филогенетическая история осевой мускулатуры позвоночных, мускулатуры конечностей).
История органов чувств показывает их постепенную дифференциацию из общих покровов, а вместе с тем и обособление соответствующих нервных центров. С другой стороны, и дифферен-цировка мышечной системы сопровождается усложнением строения нервной системы. Наконец, и многие преобразования в скелете зависят от функции связанных с ними мышц. Частично все эти изменения строятся за счет функциональных модификаций. Это доказывается экспериментами выключения нормальной функции мышц или нервов, ведущими к частичному недоразвитию скелетных и мышечных образований. Многие такие изменения наблюдаются уже при содержании диких зверей в неволе. Если способность к функциональной гипертрофии в наших экспериментах и оказывается ограниченной, то вое же она настолько велика, что может обусловить неизмеримо более значительные преобразования в течение жизни одного поколения, чем самый интенсивный отбор наследственных изменений на протяжении существования многих сотен поколений.
При функциональной адаптации, связанной с установлением новых дифференцировок, эти изменения строятся, конечно, на основе наследственной, т. е. выработавшейся в предшествующей истории организма, нормы его реакций. Способность мышц к усилению в результате тренировки, способность нервных центров к некоторой гиперплазии, способность скелетных частей усиливаться и перестраиваться соответственно измененной нагрузке и все другие выражения функциональной гипертрофии основываются на унаследованной норме реакций данного организма. Она определяет и диапазон возможных изменений и, следовательно, границы возможной индивидуальной приспособляемости. Однако унаследованной оказывается лишь данная реакционная способность в общем — для мускульной ткани, для нервной ткани и для скелетных образований. Вполне, конечно, возможны различия в реакционной способности тех или иных категорий мышц (мышц конечностей, осевых или висцеральных мышц), областей нервной системы (первичных двигательных ядер и высших областей первичных и вторичных чувствующих центров) и разных скелетных структур (пластинчатые, трубчатые и компактные кости). Однако совершенно невероятным было бы предположение, что каждая отдельная мышца обладает своей строго определенной способностью к тренировке. Многие различия определяются, очевидно, только тренировкой, только реальным распределением функций в организме, находящемся в данной конкретной обстановке.
В некоторых случаях удается установить значение функции даже в развитии «пассивных» образований. Известно, что кожа млекопитающих реагирует на постоянное давление или трение образованием мозолей. Это — общая реакция, которая, однако, приобретает иногда значение постоянной местной реакции (например, подошвенные мозоли на лапах многих млекопитающих). И у человека на подошвах ног кожа значительно утолщена. Свинья-бородавочник (Phacochoerus) разрывает землю для добычи корма и при этом всегда длительно опирается на локтевой сустав. Соответственно на локтях у нее развиваются постоянные мозоли. Так как эти мозоли, очевидно, появились в результате новой привычки (а не наоборот,— нельзя допустить, что свинья стала рыть землю в такой позе именно потому, что у нее развились мозоли), то первоначально они имели характер чисто модификационных изменений, возникших на базе общей реакционной способности кожи. Здесь произошла, следовательно, явная дифференциация — на базе исторически развившейся общей реакционной способности выделилась специальная реакция местного характера (местная мозоль) как приспособление к особым условиям существования Именно это частное модификационное изменение приобрело ведущее значение в дальнейшей эволюции. В процессе эволюции оно стабилизировалось как наследственное изменение. Исследования Лехе (Leche, 1902) показали, что эти мозоли закладываются уже у зародыша. Точно так же указывают, что и у человеческого зародыша эпидермис на подошве ног оказывается уже утолщенным. Следовательно, то образование мозолей, которое возникло первоначально как модификационное изменение в виде ответа ' на местное раздражение, стало затем развиваться на соответствующем месте и без этого внешнего раздражителя, под влиянием внутренних факторов (автономизация развития). Это означает стабилизацию данного новообразования, возникшего под непосредственным дифференцирующим влиянием функционального раздражения.
г. Стабилизация конкретных модификаций. Если при переходе из одного биотопа в другой физиогенная модификация имеет скачкообразный характер замены одной хорошо выраженной адаптивной нормы совершенно иной (водная форма, ксерофитная, альпийская и т. п.), то это, конечно, не составляет общего правила. Гораздо чаще историческое изменение условий существования совершается более постепенно и сопровождается таким же постепенным приспособлением организма.
Особое значение имеют при этом общефизиологические и функциональные модификации. Они реализуются в пределах существующих общих норм реакций в виде дифференцированных частных реакций.
Стабилизация конкретных результатов таких реакций должна происходить во всех тех случаях, когда данная модификация становится общим достоянием целой популяции в ряде поколений. Мы знаем, что стабилизирующая форма отбора осуществляется на базе селекционного преимущества приспособленной (в том числе и вновь приспособленной) нормы перед всеми от нее уклонениями, т. е. путем накопления всех мутаций, не выводящих фенотип за пределы установившейся нормы.
Можно, конечно, предполагать и одновременное действие обычной формы естественного отбора в том же направлении, в каком происходит модифицирование организма (на основе селекционного преимущества дальнейших отклонений по сравнению с уже осуществленной нормой). Это возможно, если модификация недостаточна, мало устойчива (в данных условиях) или реализуется лишь поздно в онтогенезе данной формы. На такую замену модификаций наследственными изменениями, обладающими известными преимуществами, обратил внимание в особенности Лукин (1935, 1936, 1940). Я не отрицаю этих процессов. Однако это не есть особая форма естественного отбора. Поскольку отбор основывается в этом случае на положительных наследственных изменениях, а возможность таких изменений довольно ограничена, то это, очевидно, весьма медленный процесс (ведущая форма отоора). С другой стороны, стабилизирующий отбор не идет далее установившейся нормы и основывается не на положительных уклонениях, а на «нейтральных» изменениях (т. е. не только на безразличных, но и на частично и условно вредных мутациях, вредные выражения которых нейтрализуются; главным же образом вообще на малых мутациях). Эти мутации встречаются неизмеримо чаще и, главное, свободно размножаются, накапливаются и комбинируются в популяциях, в пределах нормального «дикого» типа. В мобилизационном резерве внутривидовой изменчивости имеется всегда готовый запас таких мутаций и комбинаций, которые, оставаясь в пределах нормы, оказываются в данных условиях более жизнеспособными, т. е. оставляют более значительное потомство, чем любые отклонения от нормы (независимо от того, каковы источники этих уклонений). Только это и составляет основу процесса стабилизации. Особи с наследственным уклонением, реагирующие на случайные факторы внешней среды возвратом к прежней норме, утратившей свое значение, элиминируются. Этим путем размножаются более устойчивые комбинации, «фиксирующие» достигнутый уровень приспособления. На основе этой новой нормы возможны дальнейшие конкретные модификации, которые по мере приобретения ими постоянного значения (в переживании особей данной популяции) в свою очередь стабилизируются в виде дальнейшей нормы, которая может служить вновь исходной позицией для прогрессирующего приспособления. Таким образом, я вовсе не думаю, что сначала происходит модификация в полной мере ее потенциальной возможности, затем происходит ее дальнейшее совершенствование через обычный отбор и, наконец, фиксирование через стабилизирующий отбор.
Я представляю себе непрерывную кооперацию этих процессов. С одной стороны, организм непрерывно до известной степени модифицируется, и, с другой стороны, те выражения индивидуальной и, в частности, функциональной адаптации, которые уже приобрели постоянное значение в данных условиях, непрерывно стабилизируются (через элиминацию фактических уклонений). Это означает сужение нормы реакций в известном направлении, утратившем свое значение. Однако через ведущую форму отбора норма реакций меняется в иных направлениях. Устанавливаются, быть может, некоторые новые адаптации, а вместе с тем расширяется в определенном направлении, диктуемом реально встречающимися факторами внешней среды, и способность к дальнейшим адаптивным модификациям (расширение нормы реакций и лабилизация формообразования).
Континентальный климат, с господством резких колебаний влажности, инсоляции и, в особенности, температуры, должен способствовать выработке наиболее стабильных форм, обладающих наиболее высоко развитой системой регуляторных механизмов. Поэтому мы считаем весьма вероятными известные гипотезы, связывающие эволюцию покрытосеменных растений, с одной стороны, и млекопитающих животных, с другой, с засушливыми периодами мезозоя и с континентальными климатическими зонами земной поверхности.
4. ВОЗНИКНОВЕНИЕ РЕГУЛЯТОРНЫХ МЕХАНИЗМОВ ФОРМООБРАЗОВАНИЯ (ИНТЕГРАЦИЯ)
В предшествующем изложении мы много говорили о стабилизации форм, об устойчивости формообразования, связанной с aвтономизацией развития, о защитных и регуляторных механизмах индивидуального развития. Теперь нам надлежит остановиться на источниках и путях развития этих механизмов, обеспечивающих осуществление нормы при изменении внешних, а частично и внутренних факторов развития. При этом мы будем отчасти исходить из наличия существующих уже физиологических связей общего значения, в основном же будем опираться на те выражения взаимосвязей, которые проявляются в отдельных мутациях (плейотропия) или создаются в процессах их комбинирования (в сбалансированных генетических системах). Мы начнем с последних, так как именно в них создаются совершенно новые системы связей, которые в процессе дальнейшей эволюции могут послужить основой для их углубления и преобразования в морфогенетические и физиологические взаимозависимости.
А. ГЕНЕТИЧЕСКИЕ СИСТЕМЫ
Прогрессивная эволюция организмов протекает в популяциях скрещивающихся между собой, генетически всегда разнородных особей и основана на естественном отборе наиболее приспособленных (на всех стадиях развития) индивидуальностей. Непрерывное перекомбинирование под контролем естественного отбора ведет к выделению наиболее жизненных генетических сочетаний (вершины приспособления или «адаптивные пики» С. Райта). В этих сочетаниях доминирующая роль принадлежит всегда малым «физиологическим» мутациям. В генетическом отношении наиболее жизненные особи являются наиболее «сбалансированными генетическими системами». Эффект таких комбинаций, как мы знаем, не является суммарным; он имеет свою качественную характеристику. В данном биотопе и данной популяции преобладающее значение приобретает некоторая общая генетическая основа, видоизменяемая лишь в деталях. Постоянная кооперация «нормальных» для данной популяции сочетаний факторов ведет ко все большей их слаженности в процессах развития. Нейтральные, вначале просто «сопутствующие» факторы, участвуя в общих процессах обмена, входят мало-помалу все глубже в обший механизм индивидуального развития и становятся необходимыми его факторами. Кладется начало обязательным сочетаниям, в которых одни факторы становятся зависимыми от других. Такое установление генетических зависимостей наблюдается и на крупных мутациях (дополнительные или «комплементарные» факторы), однако несомненно гораздо большее значение имеют целые комплексы малых мутаций и генов-модификаторов. В процессе естественного отбора наиболее благоприятных в данных условиях выражений отдельных мутаций создаются такие оптимальные сочетания модификаторов, непрерывно связанные с данными мутациями в целостные комплексы. Создаются сбалансированные генетические системы, которые могут иметь ограниченное распространение и как локальные формы вести к дальнейшему их расхождению, но на равных основаниях могут установиться в одной и той же популяции и при скрещиваниях давать картину альтернативного наследования целых систем (типичный гетероморфизм).
Внесение чуждых элементов путем скрещивания с другими локальными формами может привести к частичному разрушению установившейся системы, к резкому возрастанию изменчивости и к появлению многочисленных мало жизнеспособных гибридных особей (полулетальный и летальный эффекты комбинаций с нарушенным генным балансом). Несовместимость генетических систем сказывается иногда и при скрещивании отдельных мутаций. Наконец, нередко даже отдельная мутация, т. е. изменение одного «гена», ведет к такому же нарушению баланса, и тогда и отдельная мутация оказывается летальной или полулетальной.
Если бы, однако, не существовало механизмов, поддерживающих баланс генетических систем, то вообще любая мутация и любое скрещивание должны были бы разрушать эти системы. Неустойчивость органических форм была бы столь велика, что не допускала ни скрещивания, ни мутирования.
Этого нет благодаря тому, что генетические системы устанавливаются при некотором ограничении панмиксии, обладают своими средствами связи, поддерживающими их целостность, имеют всегда, до известной степени, регуляторный характер и, кроме того, дополняются морфофизиологическими регуляторными механизмами, которые приобретают в процессе прогрессивной эволюции постепенно все большее значение.
Установлению определенных генетических комбинаций способствует прежде всего известное ограничение свободы скрещивания, создаваемое различными формами изоляции (см. II—8, В). Затем известную роль играют, вероятно, явления сцепления. Воз-можно, что в самих хромосомах мы имеем первичное средство связи, помогающее установлению сбалансированных комбинаций в пределах каждой хромосомы. Все это, однако, средства, имеющие лишь некоторое значение на начальных этапах формирования новых биотипов. Гораздо большее значение должны иметь регуляторные механизмы. В качестве таковых мы рассматриваем явления диплоидности, генный баланс (геномные корреляции регуляторного характера), доминантность нормы и морфофизиологические регуляции (феногенетические, морфогенетические и эргонтические корреляции).
Диплоидность. Разрушающее влияние отдельных мутаций ослабляется или даже вовсе устраняется вследствие диплоидной организации ядра, при которой любому мутантному гену всегда противостоит его нормальный аллеломорф. Поскольку все мутации распространяются, комбинируются и отбираются только в гетерозиготном состоянии, каждая особь известной популяции содержит всегда полный комплекс нормальных аллеломорфов (в обоих геномах), т. е. одну полную сбалансированную генетическую систему, а сверх этого значительную часть второй такой же системы. На этом вполне устойчивом фоне выделяется лишь относительно небольшое число мутантных генов, за счет которых создаются все новые комбинации, включаемые в нормальную генетическую систему лишь по мере полного установления нового баланса.
Генный баланс. Регуляторный характер сбалансированных генетических систем выражается уже в том, что мутации, обладающие ясным выражением в одной системе, могут полностью потерять это выражение в другой системе. О том же говорит самое существование мутаций с неполным проявлением, так как эти различия, которые частично могут быть объяснены случайными внешними влияниями, в основном покоятся все же на малых генетических различиях отдельных особей, всегда имеющихся даже в чистой культуре изогенной линии. Это заставляет думать, что некоторые мутации вообще не получают никакого выражения, т. е не проявляются вследствие того, что они не могут нарушить данную генетическую систему, достаточно защищенную регуляторным механизмом, противодействующим влиянию данных мутаций. Мы, к сожалению, не может сказать, в чем заключается генный баланс и на чем основан его регуляторный характер. Так как эти явления все же явно отличны от далее упоминаемых морфогенетических регуляций, то можно лишь строить предположения об особых формах биохимической устойчивости внутриклеточной системы, находящейся, быть может, в состоянии подвижного равновесия или обладающей своеобразными «буферными» механизмами защиты. Эта устойчивость выражается, очевидно, в явлениях внутриклеточного обмена веществ, лежащих в основе всех морфогенетических процессов.
Доминантность. Баланс нормального генотипа обычно достаточно защищен от нарушений со стороны небольших мутаций. В гетерозиготе это выражается в доминировании нормы.
Так как многочисленные мутации распространяются в популяциях лишь в гетерозиготном состоянии, то практически в борьбе за существование конкурируют всегда самые разнообразные гетерозиготы. В естественном отборе преимущества в большинстве случаев окажутся на стороне уже приспособленной «нормы». Погашение выражения различных неблагоприятных мутаций в гетеро-зиготе, т. е. усиление доминирования «нормы», достигается, как мы видели, отбором наиболее благоприятных генных сочетаний. В результате создается известная защита нормального формообразования от возможных нарушений, создается известный «фактор безопасности» (Haldane, 1929). Генетики отмечают, что в этом случае активность нормального «гена» достигает оптимального уровня, так что его удвоение в гомозиготе уже не вносит дальнейших изменений. Мы уже рассматривали факты обезвреживания мутаций в гетерозиготе (через развитие их рецессивности), а тем самым разобрали и вопрос об эволюции доминантности нормы. Мы видели, что различные авторы представляют себе эти процессы по-разному, и пришли к выводу, что в основе этих разногласий лежат вполне объективные различия конкретных путей, при помощи которых в эволюции достигаются сходные результаты. Доминирование нормы представляет во всяком случае побочный результат развития более устойчивых форм. Однако на основе этой устойчивости нормального формообразования лежат не только генетические механизмы, но и более сложные механизмы индивидуального развития, имеющие также до известной степени регуляторный характер.
Б. ФЕНОГЕНЕТИЧЕСКИЕ1 СИСТЕМЫ
1 В личном экземпляре И. И. Шмальгаузена — «эпигенетические» (Waddington).— Ред.
Основным источником преобразования системы взаимозависимостей частей развивающегося организма служат, очевидно, элементарные изменения взаимосвязи, обнаруживаемые в отдельных мутациях. Они выражаются в известных явлениях плейотропизма. Общая характеристика явлений была дана ранее (см. 1–4, Б).
Первично, очевидно, любое наследственное изменение должно было изменять весь организм в целом. Если те или иные части этого организма находятся в разном положении относительно факторов среды, т. е. процессы обмена в них идут несколько различно, то, очевидно, изменение одного наследственного фактора приведет к различным изменениям в этих, разных по своей физиологической ситуации, частях. Если эти изменения имеют, при данных условиях, необратимый для особи характер, т. е. касаются образования более стойких продуктов, то возможна на этом пути наследственная дифференциация организма, определяемая, однако, в своей реализации, на известной стадии развития, некоторыми факторами внешней среды. Дифференциация возможна и в том случае, если детерминирующие ее продукты остаются внутри тех клеток, специфический характер обмена которых определяет специфику действия данных продуктов.
По мере усложнения строения организма происходит известная дифференциация детерминирующих продуктов. Каждый из этих продуктов действует различно на различные части, но их действие всегда ограничивается теми клетками, в которых эти продукты образуются.
Эту форму плейотропного действия можно обозначить как множественное, или полиморфное, выражение мутации. Изменение одного гена сказывается так или иначе на всех клетках организма. В зависимости от уже имеющейся в наличии дифференцировки, от положения и, следовательно, характера обмена в разных клетках изменение преобретает разный характер и ведет к различным изменениям продуктов дифференцировки. Взаимозависимости в изменениях различных частей, определяемые здесь изменениями в первичных же морфогенных продуктах, мы обозначаем как геномные и, в частности, феногенетические корреляции.
Однако совершенно невероятно, чтобы изменение одного гена, ведущее к благоприятному изменению в одной части, привело к благоприятному изменению в другой части (А. А. Малиновский). Следовательно, эта форма плейотропизма, характерная для мозаичного организма, но широко распространенная и при регуляционном типе развития, сама по себе не может приобрести положительного значения в процессе эволюции. Путем непрерывного отбора других, особенно малых, мутаций происходит такое их комбинирование, которое приводит к погашению всех неблагоприятных прояв-. лений плейотропизма, т. е. к уничтожению большинства видимых выражений данной мутации. Так как, кроме одного основного проявления (представляющего собой предмет отбора), все другие, как правило, оказываются неблагоприятными, то эволюция идет все время в сторону устранения плейотропного выражения мутаций и выработки их специфически локального действия, на основе взаимодействия между различными внутриклеточными процессами, зависящими от изменения многих генов. Лишь в редких случаях, в результате отбора благоприятных комбинаций, возможно установление нескольких положительных выражений одной мутации (в сочетании с рядом модификаторов). Как правило, положительное значение в процессе эволюции могут приобрести лишь взаимозависимости, определяемые целым комплексом таких изменений. Такие «морфогенетические» корреляции выражаются уже не в явлениях обычного плейотропизма и методами генетического анализа не могут быть вскрыты.
Наиболее резко выражено локальное действие морфогенных продуктов у мозаичных организмов и, в частности, у дрозофилы. Что это действие в самом деле ограничивается теми клетками, в которых эти продукты образуются, видно по мозаикам и отдельным соматическим мутациям, в которых изменение сказывается в резко ограниченной области, простирающейся иногда (IB случае позднего возникновения) на очень небольшой участок — при изменении окраски глаза, например, на несколько фасеток.
Возможна, однако, и другая форма плейотропизма, которую можно обозначить как многостепенное или полифазное выражение мутации. В этом случае морфогенные продукты распространяются путем нередко весьма ограниченной, но иногда и более свободной диффузии, а специфика морфогенетической реакции зависит не только от этих продуктов, но и от условий их распространения в теле, а также, конечно, в значительной мере и от уровня диф-ференцировки, положения и, следовательно, характера обмена в клетках различных частей развивающегося организма. Число возможных дифференцировок, достигаемых в результате действия одних и тех же морфогенных продуктов, при этом значительно возрастает вследствие увеличения числа комбинаций действия различных веществ, достигающих разной концентрации в тканях различной ситуации, разного возраста и дифференцировки (а следовательно и обмена). Такие взаимозависимости можно рассматривать как элементарные формы морфогенетических корреляций.
Диффундирование морфогенных продуктов (в виде «геногормонов») доказано в некоторых случаях и для мозаичных организмов. Однако гораздо большее значение оно приобретает при регуляционном типе развития, как это в особенности характерно для позвоночных животных.
Уже Стертевантом (1929а) было доказано существование у дрозофилы выделяемого яичником гормона, который подавляет в нор-, мальной мухе проявление окраски vermilion. To же самое было затем подтверждено опытами Бидла и Эфрусси (1935–1937) с трансплантацией имагинальных дисков глаз, взятых от личинок различного наследственного строения, на личинки иного генотипа. Почти все глазные окраски развивались автономно, т. е. под контролем факторов, заключающихся в клетках самого зачатка. Однако окраски vermilion и cinnabar развивались в зависимости от хозяина и, следовательно, в теле нормального хозяина (и большинства других мутантов) не проявлялись. Опыты трансплантации показали также возможность обратного влияния трансплантата на ткани мутантного хозяина. Наконец, опытами инъекции лимфы показано, что действительно вещества порядка «геногормонов» распространяются в этом случае путем диффузии по всему телу (Эфрусси, Кланси и Бидл, 1936). То же самое показывают и опыты кормления личинок дрозофилы одного генотипа кашицей из личинок или куколок другого генотипа, содержащих такие диффундирующие вещества, действующие на окраску глаз (Бидл, Эфрусси, 1936, 1938; Нейгауз, 1939, 1940). Кроме яичника, местом образования этих «геногормонов» оказались также жировое тело и мальпигиевы сосуды.
Очень ясную картину гормонального влияния дали исследования Кюна и его сотрудников над развитием амбарной огнёвки Ephestia kuhniella. Здесь показана особо сложная цепь зависимостей пигментообразования в разных частях тела и в разных возрастах. Изменения вызываются и здесь «геногормонами», вырабатываемыми главным образом в половых железах, именно в семеннике (а отчасти и в центральной нервной системе), откуда они распространяются по всему телу. Изменение сводится в основном к изменению скорости пигментообразования и сказывается на окраске покровов гусеницы, ее ложных глазков, оптического ганглия и на глазах взрослой моли.
У позвоночных формообразовательное значение гормонов общеизвестно. Изучено также значение продуктов, действующих на небольшие расстояния и известных под названием индукторов (в широком смысле слова). Связь с отдельными «генами» здесь в большинстве случаев не установлена, так как не производилось сравнительного исследования действия индукторов у форм, отличающихся одним или немногими «генами». Исключение составляют лишь немногие работы, как, например, феногенети-ческий анализ куроперости, проведенный Пеннетом и Бейли (1921), феногенетический анализ карликовости у мышей по исследованиям Смита и Мак-Доуелла (1931). В первом случае изменение зависит от изменения реактивности тканей на женский половой гормон, во втором случае — от изменения продукции ростового гормона в гипофизе. К этому можно еще добавить исследования Бонневи (1934) над аномальными мышами Литтля и Багга. Здесь причиной развивающихся дефектов оказалось действие цереброспинальной жидкости, в избытке выступавшей из IV желудочка мозга и собиравшейся в различных местах под кожей, в особенности у глаз или в конечностях.
Я проанализировал у кур развитие некоторых расовых признаков, имеющих характер простых мутаций, т. е. зависящих в своей реализации от изменений в отдельных генах. Приведу несколько примеров расовых признаков со связями возрастающей сложности.
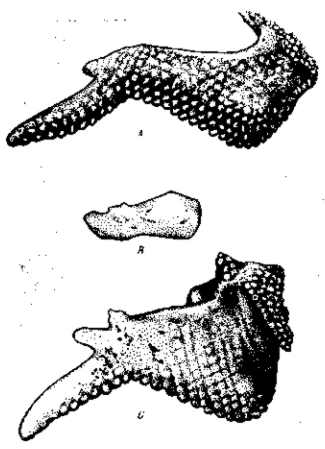
Рис. 34. Закладка лишнего пальца в крыле зародыша полидактильной курицы (фавероль) и ее регуляция. А —крыло 9-дневного зародыша обыкновенной курицы; В — крыло с раздвоенным зачатком первого пальца 7-дневного зародыша фавероли; С — крыло 9-дневного зародыша фавероль. На более поздних стадиях зачаток останавливается в своем развитии, получает очень изменчивое выражение и совершенно редуцируется.
Полидактилия кур выражается в частичном удвоении зачатка конечности по преаксиальному ее краю, в зеркальном расположении, как это наблюдается и в экспериментальных ДЕойниках. Закладываются 2–3 лишних пальца, но вполне развивается лишь один добавочный палец, дифференцирующийся по типу второго пальца. Закладка происходит и в задней и в передней конечностях, но в передней ее развитие затем подавляется (рис. 34). Следовательно, плейотропный эффект этой мутации вторично уничтожается. Здесь имеется плейотропизм первого рода, т. е. типичное множественное выражение изменения гена.
Мохноногость кур выражается у эмбриона очень ранней концентрацией мезенхимы под кожей постаксиального края задней конечности, сходной с нормальной ее концентрацией в передней конечности (соответственно закладке маховых перьев). Можно думать о восстановлении ллейотропного эффекта, который в истории птиц был полностью подавлен как вредный (здесь, естественно, вспоминается гипотетический Tetrapteryx, бывший, по Бибу, переходной формой от лазающих псевдозухнй к перво-птяцам). Концентрация мезенхимы под кожей ведет к нарушению нормального расположения зачатков чешуи на цевке, а затем к закладке ряда перьев, напоминающих маховые (рис. 35). Это, видимо, вторичный эффект изменения гена. Мезенхима, идущая на построение этих закладок, имеет явно скелетогенное происхождение, и в связи с этим более или менее недоразвиваются фаланги 4-го пальца (брахидактилия), палец изгибается на тыльную сторону, и на нем недоразвивается коготь. Об этих изменениях можно говорить как о третичных выражениях мутации мохноногости (именно доминантной мохноногости типа брама и бентамок). Ясно проявляется плейотропизм второго рода, т. е. многостепенное выражение изменения гена.

Рис. 35. Развитие мохноногости у кур. А и В — закладка первого и второго ряда перьев ноги («маховых» и «кроющих»). А — конечность 11-дневного зародыша брама с почти нормальным четвертым пальцем; Б — конечность 11-дневного зародыша брама с резко выраженной брахидактилией и дорсальным изгибом четвертого пальца; С — конечность 11-дневного зародыша бентамки с сильно развитым оперением и резко выраженной брахидактилией (вид с тыльной стороны); Д — конечность 11-дневного зародыша фавероли с зачатками перьев и вполне нормальным четвертым пальцем
Еще более сложная цепь зависимостей обнаруживается при развитии хохла у гуданов. Здесь первое видимое выражение мутации выявляется на эмбриональной стадии у шестидневных зародышей в усиленной секреции цереброспинальной жидкости клетками эпендимы переднего мозга. Образуется масса крупных вакуолей, лежащих непосредственно под эпендимой. В результате сильно раздуваются передние желудочки головного мозга (рис. 36, 37). Мозговая жидкость, очевидно, просачивается через тонкую крышу III желудочка и действует разрушающим образом на перепончатую закладку крыши черепа, полностью ее дезорганизуя (сходное действие цереброспинальной жидкости установлено и в упомянутых исследованиях Бонневи; кроме того, продукция фибринолитических ферментов клетками эпендимы показана Вейссом и в культурах in vitro). Эта дезорганизация зачатка крыши черепа является вторичным выражением хохлатости гуданов. В связи с изменением формы головного мозга стоит своеобразное перемещение зачатков костей черепа к его основанию как ряд третичных изменений (рис. 41, 42). Скелетогенная мезенхима крыши черепа концентрируется под кожей, здесь же разрастается обильная сеть мозговых кровеносных сосудов, и на этом месте развивается масса увеличенных зачатков перьев, образующих затем характерный хохол на голове. Все это можно обозначить как дальнейший ряд третичных и четвертичных изменений. Наконец уже постэмбрионально под кожей развиваются отдельные островки костной ткани, соединяющиеся затем перекладинами и образующие решетчатую вначале, своеобразную структуру— новую, сильно вздутую крышу черепной коробки (рис. 38).
Рис. 36. Поперечные разрезы через переднелобную область головы 10-дневных зародышей нормальной курицы (фавероль) и гидроцефальной (гудан). У гидроцефального зародыша желудочки переднего мозга раздуты, крыша перепончатого черепа разрушена, закладки лобных костей сдвинуты к основанию мозга fr — лобные кости, М — зачаток твердой мозговой оболочки
Рис. 37. Поперечные разрезы через лобную часть головы 10-дневных зародышей в области монроевых отверстий мозга. Вверху — фавероль, внизу — гудан. У гидроцефального зародыша желудочки раздуты, лобные кости сдвинуты к основанию переднего мозга (их дорсальные края как будто срезаны). На вздутой верхней поверхности головы видны сильно развитые зачатки перьев (хохол)

Рис. 38. Череп гидроцефальной курицы с решетчатой шишкой в лобной области и для сравнения (внизу) — череп обыкновенной курицы
Поскольку вся эта цепь изменений нормального развития зависит от изменений одного гена, мы здесь можем определенно говорить о плейотропном выражении мутации. Однако этот плейотропизм имеет явно иной характер, чем то, что мы находим у дрозофилы. Если мы там говорили о множественном эффекте, понимая под этим только различную локализацию отдельных изменений, то здесь мы можем говорить о многостепенном, или полифазном, выражении плейотропизма. Несомненно, что и здесь отдельные неблагоприятные выражения могут быть вторично подавлены действием модификаторов. Это доказывается фактом существования хохлатых кур без черепной шишки, а также исследованиями Фишера, установившего, что у японских шелковых кур выражение мутации гидроцефалии подавляется комплексом других факторов, между тем как развитие хохла остается.
Учитывая, что при многостепенном выражении изменения одного гена мы имеем явные зависимости между морфогенетическими процессами, мы можем уже с полным правом говорить о морфогенетических корреляциях.
Многостепенное выражение одной плейотропной мутации можно себе представить как результат последовательной реализации известных реакций. Если первичные морфогенные продукты образовались в самих клетках и их действие сказалось, например, в усилении секреции клеток эпендимы, то эти вторичные продукты секреции диффундировали в ткани и разрушали зачаток крыши черепа, оказав какое-то действие на составлявшие его мезен-химные клетки. Изменение в обмене мезенхимных клеток было связано с изменением их реактивной способности — они переместились под кожу. Выделяемые ими третичные продукты вызвали здесь разрастание кровеносных сосудов, более мощное развитие зачатков перьев и т. д. Это действие можно выразить диаграммой (рис. 39).
Если мы, однако, уже раньше отмечали полную невероятность того, что различные выражения изменения одного гена могут иметь положительное значение для жизни организма в какой-либо конкретной обстановке, то и для этих форм плейотропиз-ма все сказанное сохраняет свою силу. Если одно выражение изменения гена является благоприятным, то другие его выражения скорее всего окажутся неблагоприятными. Эти последние могут быть подавлены в процессе естественного отбора модификаторов. Это видно из следующего. Вторичные результаты изменения гена обычно весьма изменчивы, что само уже указывает на существование модификаторов. Так, например, мутация мохионогости выражается (в зависимости от времени мобилизации скелетной мезенхимы) весьма различной степенью брахидактилии, и в некоторых случаях это последнее выражение может быть полностью подавлено (как, например, у фаверолей). Если в процессе эволюции мохноногость может приобрести положительное значение, то побочное, отрицательное выражение той же мутации — брахидактилия, очевидно, будет подавлено.
Мутация полидактилии выражается в закладке лишнего пальца в обеих парах конечностей, но в крыле ее проявление затем подавляется. При этом изменчивость зачатка лишнего пальца крыла очень велика, что указывает на роль модификаторов в его редукции.
Мутация хохлатости кур могла бы приобрести в процессе эволюции положительное значение (хотя бы как распознавательный признак), но недоразвитие крыши черепа есть во всяком случае очень вредное изменение. И это выражение может быть подавлено, как показывает пример японских шелковых кур. Очевидно, это изменение произошло за счет уменьшения секреции мозговой жидкости и уменьшения реактивности мезенхимы, либо, еще вероятнее, за счет более раннего и свободного выделения мозговой жидкости наружу (чем предотвращалось раздувание желудочков и последующее весьма интенсивное истечение жидкости).
Таким образом, многостепенная форма выражения плейотро-пизма в процессе эволюции неизбежно осложняется изменением морфогенных продуктов, в связи с изменением других генов, что частью осуществляется через более сложные формы взаимодействия с другими тканями (т. е. через вторичные и третичные продукты). Это осложнение морфогенетических процессов может быть выражено приведенной второй диаграммой (рис. 39). Здесь вторичные и третичные (В, С, D) выражения основной мутации видоизменены взаимодействием со вторичными продуктами других клеток, которые нейтрализуют вредные выражения и, быть может, дают новые, благоприятные для жизни организма выражения комплексного действия ряда мутаций.
Рис. 39. Схема многостепенного выражения плейотропии (I), изменяемого в процессе эволюции вследствие естественного отбора модификаторов (II) и дальнейшего усложнения форм взаимодействия (III), в результате которого развивается типичная система морфогенетических корреляций
Пока мы имеем дело с цепью реакций, определяемых одной основной мутацией, вторичные выражения которой видоизменены действием модификаторов, мы с полным правом говорим еще о явлениях плейотропизма. Вместе с тем здесь, однако, совершенно ясно выступают морфогенетические корреляции между отдельными процессами, определяемыми первичными или вторичными морфогенными продуктами. Эти корреляции мы рассматриваем как элементарные формы морфогенетических корреляций, так как в процессе эволюции они всегда претерпевают дальнейшие преобразования и усложнения, в результате которых генетический анализ взаимозависимостей становится практически невозможным. В процессе исторического развития организмов изменения характера мутаций, подобные разобранным, являются лишь базой, на которой строятся видовые различия более крупных таксономических подразделений. В этом случае мы никогда не имеем дела с признаками, определяемыми изменениями единичных генов, а всегда с результатом преобразования очень многих генов, т. е. мы имеем множественную зависимость любого признака и любой фазы его развития. Это может быть пояснено третьей, еще более сложной диаграммой (рис. 39).
Естественно, что система зависимостей, каждое звено которой определяется очень многими факторами, отличается значительно большей устойчивостью, чем разобранные нами ранее выражения простого или многостепенного плейотропизма. Признаки, зависящие в своем развитии от такой сложной системы морфогенетических корреляций (как это характерно для позвоночных), отличаются значительно большей устойчивостью, чем автономные признаки мозаичного организма (например, дрозофилы).
Нормальное развитие всего организма обусловливается при наличии сложной системы корреляций всем генотипом в целом. Роль отдельных генов снижается в связанных частях организма до уровня модификаторов. Система корреляций обеспечивает, таким образом, устойчивость основных черт организации, а возможная здесь без летальных последствий высокая мутабиль-ность отдельных генов гарантирует достаточную пластичность конкретного ее оформления.
В. МОРФОФИЗИОЛОГИЧЕСКИЕ СИСТЕМЫ
По море усложнения системы корреляций, последние теряют свой «генетический» характер, т. е. становятся практически неразложимыми (точнее — их нарушение в результате мутации приводит к летальным последствиям). В таком случае можно говорить о морфофизиологических корреляциях и их системах, как единицах, не поддающихся обычному генетическому анализу. Эти системы корреляций составляются в своей основе из взаимозависимостей в жизненных функциях организма, благодаря которым происходит согласованное течение общефизиологических процессов. В первую очередь это касается клеточного обмена веществ, связанного с образованием морфогенных веществ в развивающемся организме. Затем, это предполагает согласование специфических свойств этих морфогенных веществ с реактивной способностью тканей на различных стадиях развития, т. е. детерминацию и регуляцию формообразования организма (морфогене-тические корреляции), и, наконец, это означает согласование общежизненных функций отдельных частей и органов развивающегося эмбриона, личинки и сформировавшегося организма (собственно физиологические корреляции). Типично физиологические взаимозависимости определяют течение различных жизненно важных процессов, которые в общем характеризуются своей легкой и быстрой обратимостью. Эти взаимозависимости имеют, как правило, регуляторный характер. Их значения мы здесь не рассматриваем, а ограничиваемся морфологическими выражениями зависимостей.
Однако и физиологические зависимости могут обладать своим морфологическим выражением. Это касается некоторых общефизиологических процессов, связанных с ростом и размножением организма, и многих функциональных взаимозависимостей, связанных с побочным морфогенетическим эффектом в виде менее легко обратимых изменений формы и структуры. Это прежде всего взаимное трофическое влияние органов, находящихся в функциональной взаимосвязи. Результатом может быть усиленный рост или ускоренная дифференцировка этих органов («функциональная гипертрофия»).
Это трофическое влияние может частично перейти в детерминирующее влияние, если в результате активации функциональных взаимосвязей происходит развитие тех или иных специфических гистологических или общеморфологических структур (например, структура и форма кости под влиянием деятельности мышц). В этом случае я говорю об эргонтических корреляциях (от греческого эргон — работа). Здесь морфогенетический эффект является, так сказать, побочным результатом существования функциональной взаимозависимости.
Наконец, взаимодействие частей в развивающемся организме может вести уже прямо к формообразовательным процессам. Формообразование является тогда не побочным результатом существования взаимозависимости между двумя или более частями, а основным ее выражением. В этом случае я говорю о морфоге-нетических корреляциях.
Ясно, что между физиологическими, эргонтическими и морфо-генетическими корреляциями имеются все переходы. У взрослого организма формообразовательные процессы сведены к минимуму и взаимосвязь частей выражается в форме физиологических корреляций между легко обратимыми жизненными процессами. У молодого, еще развивающегося организма, лишь вступающего в пору своей дефинитивной жизнедеятельности, эти (физиологические корреляции играют в то же время и роль факторов, обусловливающих развитие многих так называемых функциональных структур, которые дают окончательную отшлифовку развивающимся формам, т. е. выступают в роли эргонтических корреляций. У эмбриона же, у которого почти вся энергия расходуется на формообразовательные процессы, подобные корреляции обусловливают взаимозависимость между этими, характерными для эмбриона, формами жизнедеятельности. Здесь корреляции выступают в роли специфических морфогенетических взаимозависимостей.
Резких граней между этими формами корреляций провести нельзя — они переходят одни в другие, меняя свое выражение по мере развития самого организма. Нетрудно себе также представить и процесс смены форм корреляций во время филогенетического развития организмов, как, например, постепенное замещение некоторых эргонтических корреляций морфогенетиче-скими, по мере «эмбрионализации» самого индивидуального развития.
Вопрос о возникновении новых морфогенетичеоких корреляций не представляет особых трудностей, если к нему подходить с точки зрения морфолога. В процессе эволюции, одновременно с новыми дифференцировками, обязательно устанавливаются и новые взаимозависимости как внутренние факторы, определяющие наследственное осуществление этих дифференцировок (т. е. процессы интеграции являются условием, без которого невозможна и наследственно детерминированная дифференциация). Система корреляций, связывающих развивающийся организм в одно целое, имеет, следовательно, ту же историческую основу, что и сама организация. Поэтому исторически сложившиеся взаимоотношения частей и являются вместе с тем на каждой стадии развития в роли факторов, определяющих дальнейшее течение процессов развития.
Между всеми так или иначе связанными частями обнаруживаются взаимодействия, которые в большей или меньшей мере приобретают детерминирующее значение. Таким образом, типичные морфогенетические корреляции развиваются всегда на базе исторически сложившихся соотношений между частями развивающегося организма. При этом, однако, если установление новых корреляций связано с возникновением любой новой дифферен-цировки, то разрушение старых корреляций неизменно сопровождает редукцию органов, потерявших свое значение.
Г. РАЗВИТИЕ МОРФОГЕНЕТИЧЕСКИХ РЕГУЛЯЦИЙ
Морфогенетические регуляции имеют, конечно, физиологическую, а в конечном счете — именно биохимическую основу. Все же мы их выделяем, поскольку они связаны с защитой нормального формообразования (возникновения собственно физиологических регуляций — огромной области эволюционной физиологии — мы здесь не касаемся). Такой защитой являются уже рассмотренные генетические системы, сформирование которых означает, очевидно, установление более устойчивых форм биохимического равновесия в процессах внутриклеточного обмена и синтеза живого вещества. Целостность таких сбалансированных систем обеспечивается вначале частичной изоляцией отдельных популяций, сложностью самих сочетаний, не допускающих их нарушения (без летальных последствий), диплоидностыо организмов, что гарантирует наличие по меньшей мере, одной полной «нормальной», генетической системы (в двух геномах) в любой почти особи природных популяций, и доминантностью нормальных аллеломорфов. Эти генетические системы защищают до известной степени развитие нормы. Однако не следует думать, что «баланс» представляет особую форму равновесия между «генами». Термины генетиков, например, «генный баланс», доминантные «гены», двойной запас «активности генов» и т. п., служат лишь упрощенными выражениями, удобными для описания сложных явлений. За этими терминами скрываются, однако, очень сложные процессы взаимодействия не «генов», а в первую очередь процессов внутриклеточного обмена веществ, и во вторую очередь — более сложных физиологических и, наконец, морфогенетических процессов. Поэтому все формы «защиты» нормального формообразования, все формы морфогенетических регуляций являются всегда результатом взаимодействия между физиологическими процессами и выражением взаимозависимости частей в корреляционных системах развивающегося организма. Если мы выделяем все же генетические и мор-фофизиологические системы, то это отражает процесс усложнения форм взаимозависимостей, которые в начале своего появления поддаются методам генетического анализа, а в дальнейшем процессе эволюции становятся уже неразложимыми при помощи этих методов (как это было только что показано).
Это усложнение ведет и к дальнейшему увеличению стойкости основ организации вследствие установления комплексного характера всех наиболее ответственных морфогенетических процессов и вследствие приобретения ими все более ясно выраженного ре-гуляторного характера. Путь развития таких регуляций, очевидно, сходен с рассмотренными уже процессами установления авторегуляции зависимых (от внешних факторов) форм индивидуального развития (см. I—5 и III—3, В).
Первоначальное действие «геногормонов» или индуцирующих веществ на известные ткани можно себе представить как влияние, нарастающее в своей интенсивности пропорционально концентрации веществ, доходящих до этих тканей (конечно, в известных пределах — от минимума, оказывающего вообще какое-либо влияние, до максимума, дающего наиболее значительную реакцию).
В процессе эволюции, вместе с изменением всего генотипа (и процессов внутриклеточного обмена) изменяются и качество и концентрация морфогенных веществ («активность» генов), а также и реактивность отдельных тканей. Концентрация морфогенных веществ, доходящих до реагирующих тканей, поднимается до уровня, обеспечивающего наступление реакции не только при уклонениях в факторах внешней среды (меняющих их продукцию или реактивность тканей), но и при мутациях. Это последнее достигается удвоением «нормальной» концентрации, дающей полный эффект в гомозиготе. Тогда проявляется доминирование нормы, т. е. нормальная концентрация морфогенных веществ и нормальное формообразование уже у гетерозиготы. Это означает вместе с тем и ограничение реактивности тканей, которые дают одну и ту же реакцию при простой (А) и при двойной (2А) дозе раздражителя. Однако новая мутация может означать не только уменьшение концентрации более чем вдвое (поэтому новые мутации часто полудоминантны), но и качественное изменение веществ. Она может означать и изменение реактивности тканей.
Поэтому нормальная реактивность тканей «охраняется» также известным «запасом», лежащим за пределами обычных колебаний концентрации морфогенных веществ (раздвигание обоих порогов реактивности). Кроме того, она «защищается» расширением этой реактивности и от качественных изменений тех же веществ (снижение специфичности индуцирующих веществ) и, наконец, «защищается» самой сложностью всего комплекса взаимодействующих факторов (зависимостью реактивности тканей и специфики морфогенных веществ от всего генотипа в целом и сложностью морфогенетических взаимозависимостей).
а. Раздвигание порогов нормальной реактивности тканей. Ес-тественый отбор наиболее жизнеспособных особей означает выработку оптимальной реактивности тканей при существующих обычных условиях поступления морфогенных веществ.
Последнее может подлежать некоторой изменчивости, зависящей как от мутаций, так и от уклонений в условиях развития. Весьма важно, чтобы оптимальная формообразовательная реакция всегда осуществлялась независимо от этих уклонений. Это и достигается в процессе эволюции путем стабилизирующего отбора (т. е. через элиминацию особей с уклонениями от нормальной формообразовательной реакции). Морфюгенетический эффект оказывается нормальным, почти независимо от концентрации морфогенных или индуцирующих веществ, по крайней мере в известных пределах обычно встречающихся уклонений от «нормальной» концентрации. Это ясно видно по материалам, изученным Гольд-шмидтом, в особенности по интерсексам непарного шелкопряда, а также и на примерах развития зависимых вторичнополовых признаков позвоночных.
Вырабатывается нижний, а быть может, и верхний порог концентрации веществ, необходимой для наступления нормальной (т. е. оптимальной) реакции (рис. 40). Чем шире раздвинуты оба порога, тем более развитие данной части оказывается обеспеченным авторегуляцией, как можно назвать это явление. Нормальное развитие обеспечено тогда и при мутациях, меняющих интенсивность образования морфогенных веществ в известных, конечно, пределах (особенно в сторону увеличения).
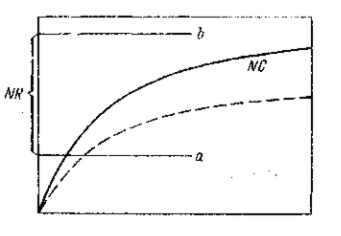
Рис. 40. Проявление известной нормы реактивности ткани. Развитие нормы стабилизируется через установление нижнего и верхнего порогов концентрации веществ, необходимой для наступления нормальной реакции. Морфогенные вещества теряют при этом свое детерминирующее значение и становятся в основном активирующими или освобождающими факторами. На оси абсцисс показано течение времени, а на ординате — нарастание концентрации веществ а — нижний и b — верхний порог нормальной реактивности ткани (NR)\ NC — кривая нормального нарастания концентрации морфогенных веществ. Прерывистой линией показано значительное снижение продукции морфогенных веществ, которая остается, однако, в пределах нормальной реактивности ткани. Такое изменение (мутация) имеет скрытые формы, но приближает организм к нижнему порогу нормальной реактивности и, следовательно, снижает устойчивость нормы, делает ее более доступной для выявления следующей мутации (или модификации), лежащей в том же направлении
Однако в действительности индуцирующие вещества поступают иногда даже не из одного источника, а из нескольких, и тогда нормальное развитие оказывается еще более обеспеченным. В этом случае возможны гораздо более значительные изменения в отдельных индукторах без того, чтобы нормальная комплексная реакция потерпела существенный ущерб. Получается нечто вроде «множественного обеспечения» нормального развития. Однако всякое уклонение, выходящее за пределы нижнего порога, приведет все же к выпадению известной реакции и явлениям недоразвития, а уклонение, выходящее за верхние пределы, может привести к заметному уклонению от нормального развития затронутых частей. Такие крайние уклонения и выявятся сразу в форме видимых «больших» мутаций или же в виде летали, если они затрагивают жизненно важный морфогенетический процесс.
В более сложных индукционных системах, связанных целой сетью многостепенных морфогенетических корреляций, каждое звено которых зависит от многих факторов типа активаторов или гормонов, достигается в конце концов относительно очень высокая степень устойчивости развивающейся авторегуляторной системы. В каждом звене имеется соотношение, которое может быть пояснено приведенной диаграммой (рис. 41), показывающей простейшую форму взаимодействия. Взаимные сдвиги во времени созревания активатора (A) и реактора (R) не оказывают влияния на нормальное течение морфогенетической реакции, пока они оба лежат в пределах между начальным и конечным пороговыми уровнями. Однако более значительное расхождение обоих компонентов индукционной системы, когда зрелая уже ткань активатора вступает в контакт с еще не созревшей или уже перезревшей и поэтому неактивной тканью (R), приводит сразу к полному выпадению данной морфогенетической реакции, а также и всех дальнейших зависящих от нее реакций. В результате происходит либо редукция этой части, либо очень большое недоразвитие, имеющее почти всегда летальные последствия, в особенности если нарушение коснулось одного из ранних и основных морфогенети-ческих процессов или жизненно важных органов.
рис. 41. Упрощенная схема одного звена индукционной системы, состоящего из активатора (А) и реактора (R), которые изменяются во времени в направлении, показанном стрелками. Периоды зрелости обоих компонентов отмечены жирными участками линий. Система имеет регуляторный характер: она допускает в известных пределах возможность взаимных смещений во времени созревания взаимодействующих компонентов без того, чтобы реакция потерпела какой-либо ущерб.
То же самое касается и изменений в продукции активирующих веществ. Изменения, не выходящие за пределы нижнего и верхнего порогов концентрации веществ, вызывающей нормальную реакцию, не дадут никакого эффекта. При выходе же за эти пределы реакция либо вовсе не наступит (при недостаточной концентрации), либо, быть может, в некоторых случаях даст резкие уклонения от нормы (при чрезмерной концентрации). И в том и в другом случае процессы развития окажутся глубоко нарушенными и обнаружатся в виде явной мутации.
Огромное большинство мутаций практически не затронет ранних стадий развития частей, связанных системой морфогенетиче-ских корреляций. Обычные небольшие мутации могут получить видимое выражение лишь на поздних стадиях развития, когда система морфогенетических корреляций уже потеряла свое значение вследствие завершения основных процессов морфогенеза.
б. Снижение специфичности морфогенных веществ. Раздвигание обоих порогов нормальной реактивности ткани означает вместе с тем довольно широкую возможность уклонений от «нормальной» концентрации морфогенных веществ без нарушения нормального течения формообразовательного процесса. Количественные изменения морфогенных факторов в известных пределах остаются без влияния на развитие нормы. Однако то же самое устанавливется затем и но отношению к качественным изменениям. В процессах мутирования, несомненно, происходят и качественные изменения специфики морфогенных веществ, а не только количественные изменения в скорости их образования и в достигаемой концентрации. Элиминация возникающих при этом отклонений от нормы означает отбор особей, наименее реагирующих на качественные изменения морфогенных веществ. Таким образом, стабилизирующий отбор приводит к освобождению реагирующих тканей от влияния случайных изменений в концентрации и в специфических свойствах индуцирующих и вообще формативных субстанций.
Следовательно, параллельно с выработкой определенной нормальной реактивности тканей, в процессе эволюции теряют свое значение не только концентрация (в известных пределах, определяемых обоими порогами реактивности), но и специфические свойства морфогенных веществ (очевидно, также в известных пределах).
Специфика реакции определяется свойствами (генотипом) реагирующей ткани на данной стадии и в данном физиологическом положении, а значение морфогенных веществ низводится до уровня несиецифических активаторов.
Мы видели, что при авторегуляторном типе индивидуального развития факторы внешней среды теряют свое детерминирующее значение и становятся лишь раздражителями, освобождающими автономное в дальнейшем течение морфогенетичеоких процессов. Точно так же мы видим, что при прогрессивной автономизации онтогенеза внутренние морфогенные факторы теряют свою специфику и их значение снижается, наконец, до уровня раздражителей, освобождающих автономное течение специфических формообразовательных процессов («самодифференцирования»).
Все без исключения опыты обменной трансплантации зачатков, переносимых с зародышей одного вида на другой, показывают, что специфика морфогенной реакции определяется свойствами самой реагирующей ткани. Последняя образует типичные для нее дифференцировки даже и в том случае, если детерминирующее влияние исходит от индуктора, относящегося к другому виду, в котором такие дифференцировки никогда не осуществляются.
Уяче первые опыты обменной трансплантации эктодермы молодых гаструл разных видов тритона показали, что нервная трубка, развивающаяся из трансплантированной эктодермы, взятой с бока или брюха Triton cristatus, под влиянием крыши первичной кишки зародыша Т. taeniatus дифференцируется все же в точности по типу Т. cristatus. С другой стороны, эктодерма Т. taeniatus, которая, оставаясь на месте, дала бы начало обыкновенному эпидермису бока, дает после пересадки в область спины зародыша Т. cristatus под влиянием примыкающего снизу индуктора, т. е. крыши первичной кишки хозяина (Т. cristatus), начало нервной трубке. Однако эта последняя, хотя и образовалась под действием морфогенных веществ Т. cristatus, развивает все специфические особенности строения нервной системы Т. taeniatus. Подобных опытов было сделано очень много с индукцией хрусталика и других органов под влиянием чуждых индукторов. При этом были получены еще более удивительные результаты.
При трансплантации головной эктодермы зародыша аксолотля на бок головы зародыша тритона в этой эктодерме никогда не развиваются характерные для тритона опорные нити — «балансеры». Однако в обратном эксперименте пересадки туловищной эктодермы тритона на боковую часть головы зародыша аксолотля, под индуцирующим влиянием аксолотля (у которого балансеров никогда не бывает), в этой эктодерме развивались балансеры типичного для тритона строения (Harrison, 1925; Mangold, 1931). Далее производились обменные трансплантации головной эктодермы между тритонами, с одной стороны, и бесхвостыми амфибиями— с другой (Rotmann, 1935; Spemann и Schotte, 1932; Spe-raann, 1936). В эктодерме тритона развивались у зародышей жерлянок и других бесхвостых, под их индуцирующим влиянием, типичные тритоньи балансеры, а в пересаженной эктодерме бесхвостых развивались на тритонах, под их индуцирующим влиянием, типичные присоски и даже роговые челюсти головастиков.
Это показывает с полной ясностью, что видовая специфика зависимых морфологических структур определяется исключительно реагирующей тканью. Морфогенные влияния, хотя и необходимы в качестве факторов, определяющих осуществление, а также время и место наступления реакции, не определяют, однако, ее качества. Эти влияния лишены видовой специфичности.
Дальнейшие попытки проникнуть в природу морфогенных или индуцирующих веществ показали, кроме того, широкую замещае-мость одних индукторов другими и выявили возможность их замены другими тканями, в которых нельзя было и подозревать существования морфогенной активности, а также различными препаратами, в том числе и синтетическими химическими соединениями. Нервная пластинка, нормально развивающаяся под индуцирующим влиянием крыши первичной кишки (хордомезо-дермы), может быть индуцирована также всевозможными другими тканями и притом не только свежими, но и убитыми нагреванием, высушиванием или замораживанием (Holtfreter, 1932!, 1933). Оказалось даже, что некоторые ткани (эктодерма), в живбм состоянии не обладающие индуцирующими свойствами, приобретают таковые после высушивания при 60° (Holtfreter, 1933). Наконец, положительные результаты были получены с эфирными экстрактами зародышевых тканей (Waddington, Needham, 1934), с нуклео-протеидами и даже с синтетическими масляными кислотами и другими соединениями (Fischer, 1935 и др.)- Положительные результаты были получены и в опытах индукции хрусталика глаза при помощи различных тканей.
Опытным путем была получена индукция добавочных конечностей у тритонов путем пересадки в бок зародыша слухового пузырька (Balinsky, 1925). Позднее удавалось достичь того же и пересадкой обонятельных мешков, регенерационной бластемы и др. Н. Драгомиров (Dragomirov, 1929) показал, что волокнистое ядро хрусталика, нормально индуцируемое ретинальным листком глазного бокала, может быть в эксперименте индуцировано и слуховым пятном лабиринта или слуховым ганглием. Он же нашел (1936), что таким же мало специфическим влиянием контакта со слуховым пузырьком может быть индуцировано и развитие добавочной ретины в пигментном листке глазного бокала.
Следовательно, индуцирующие ткани и вещества не только лишены видовой специфичности и могут быть широко заменены соответствующими тканями и органами, взятыми от других животных, но не имеют и морфогенной специфики. В наибольшей степени это касается «первичного организатора», под влиянием которого развиваются нервная система и все осевые органы. В широкой мере неспецифичны и те влияния, которые вызывают образование добавочных конечностей. Однако здесь возможность индукции все же ограничена известной областью бока зародыша,— очевидно, здесь имеются необходимые добавочные влияния, быть может не лишенные некоторой специфичности. Кроме того, можно указать, что наиболее активным из чуждых индукторов является обонятельный мешок и именно его каналовая часть; несколько менее активен слуховой пузырек. Другие органы, очевидно, лишены этой активности. В индукции волокнистой структуры хрусталика нормальное влияние ретины может быть заменено весьма сходным, очевидно, влиянием слухового пятна или слухового ганглия, но оно не может быть заменено влиянием обонятельного мешка. Последний в этом случае оказывается недеятельным.
Таким образом, отсутствие органогенной специфики сказывается особенно ярко в индукторах наиболее общих и важных образований (осевых органов, конечностей) и гораздо менее выражено в частичной замещаемости индукторов частных структур (хрусталика и ретины глаза). Очевидно, это связано с тем, что формообразовательный аппарат более общих основ организации проделал уже более длинный путь эволюции в направлении утраты специфики морфогенетических влияний и установления максимальной автономности развития важнейших систем органов. Этим самым достигнута максимальная защита формообразования от каких-либо нарушающих влияний. Случайные изменения концентрации или химических свойств морфогенных веществ не могут тогда нарушить нормального развития основных органов животного. Это не значит, что сама «индукция» потеряла свое значение. Контакт индуктора и реактора определяет в индукционной системе если не специфику, то время наступления и, в особенности, согласованность положения и протяженность взаимозависимых морфогенных реакций и тем самым определяет возможность регуляции и сохранения гармонических соотношений между органами.
Эволюция онтогенеза идет в общем по пути прогрессивной автономизации важнейших морфогенных процессов, связанной с указанной уже регуляторной организацией индукционных систем. Эта организация допускает, кроме изменения в концентрации и в составе морфогенных веществ, также возможность более или менее значительных сдвигов во времени и в пространстве без нарушения нормального формообразования. Она имеет и в целом и в отдельных своих компонентах регуляторный характер.
Любой индифферентный еще зачаток допускает весьма далеко идущие экспериментальные вмешательства без нарушения дальнейшего формообразования. Если зачаток действительно совершенно индифферентен, то вполне понятно, что и часть зачатка, после его разделения, дает начало все же целостному образованию. Однако зачатки никогда не бывают вполне индифферентными. Они по меньшей мере обладают известной полярной структурой, некоторым «градиентом». Естественно, что и они допускают возможность различных сдвигов без нарушения нормального формообразования. И изменение интенсивности самого градиента может оказаться без заметного влияния. Таким образом, и «индифферентные» зачатки, выступающие в роли компонентов более сложных морфогенетических систем, обладают также способностью к регуляторному восстановлению своей структуры при ее нарушении.
В процессе эволюции развивается, таким образом, целая система регуляторных механизмов, «защищающих» нормальное течение процессов формообразования от всевозможных случайных нарушений. Все эти изменения идут, очевидно, под направляющим влиянием стабилизирующей формы отбора, основывающейся на постоянной элиминации особей, формообразование которых было существенно нарушено в результате случайных внешних влияний или в результате мутаций.
Д. ЗНАЧЕНИЕ РЕГУЛЯТОРНЫХ КОРРЕЛЯЦИЙ В ЭВОЛЮЦИИ
В предыдущем изложении я уже неоднократно отмечал значение регуляций как факторов, способствующих переживанию особей в разнообразных условиях существования, содействующих накоплению скрытого резерва наследственной изменчивости и поддерживающих пластичность организмов в процессе их исторического преобразования. Так как эти моменты имеют очень большое значение и последовательно вытекают из всего рассмотренного материала, то я нахожу нужным обобщить здесь все существенное, прежде чем перейти к основным выводам относительно эволюции индивидуального формообразования, которым посвящена следующая глава.
Вопрос о значении корреляций в эволюции был впервые поднят А. Н. Северцовым (1914), а затем разработан мною (1938а, 19386, 1939а, ,19396, 19406). Здесь выдвигается главным образом вопрос об их значении на низовых этапах эволюции в самой общей форме. Более специальное освещение он получил в моей книге «Организм как целое в индивидуальном и историческом развитии».
а. Физиологическая эврибионтностъ. Установление регуляторных механизмов, противодействующих нарушениям формообразования со стороны случайных изменений в факторах внешней среды, означает ограничение способности к модификационным реакциям известными пределами одной или нескольких адаптивных норм.
Авторегуляция, в нашем определении, ведет к ограничению одной или нескольких «нормальных» реакций известными пределами интенсивности внешних факторов между нижним и верхним порогами реактивности тканей. В процессе эволюции оба порога реактивности раздвигаются шире, и если они вначале охватывают лишь область обычных небольших колебаний в интен-сивностях факторов внешней среды, то в дальнейшем они распространяются на более широкие пределы встречающихся иногда случайных уклонений. Для организма это означает повышение устойчивости формообразования и прогрессивную автономизацию развития. Об автономном развитии мы говорим в том случае, когда оба порога реактивности тканей раздвинуты за пределы встречающихся различий в факторах среды. Тогда нормальная реакция наступает при всех условиях и протекает исключительно под направляющим влиянием внутренних факторов развития (которые получают доминирующее значение и при авторегуляции).
Организм внешне не реагирует на изменения в факторах среды. Это объясняется, однако, исключительно существованием внутренних регуляторных механизмов, противодействующих внешним влияниям. Вместо морфогенетических реакций приспособительного характера при автономном развитии устанавливаются физиологические регуляции, нейтрализующие вредные влияния на развитие организма. Иными словами, морфологические адаптации заменяются физиологическими.
Физиологическая приспособляемость свойственна в известной степени всем организмам. Она возникала одновременно с самой жизнью и развивалась прогрессивно вместе с усложнением жизненных процессов. Установление авторегуляторных форм индивидуального развития, происходившее также уже на заре органической жизни, означает усиление физиологической приспособляемости организма. Собственно авторегуляция нескольких адаптивных норм достигает наивысшей ступени у высших растений. У животных авторегуляция ограничивается обычно одной основной адаптивной нормой, но дополняется весьма развитой способностью к функциональным адаптациям. Авторегуляция одной нормы достигает высших форм автономного развития только у животных. Очень совершенная система физиологических адаптации и регуляций животных усложняется еще развитием адаптивных форм движения, руководимых нервной системой (таксисами, инстинктами и, наконец, высшими, сознательными проявлениями деятельности нервной системы). Физиологическая и, в частности, нервная форма индивидуальной приспособляемости достигает у позвоночных, и особенно у высших позвоночных, наибольшего совершенства.
Физиологическая приспособляемость к изменениям во внешних факторах способствует широкому расселению и завоеванию новых мест в природе еще в гораздо большей степени, чем морфологическая приспособляемость, так как она устанавливается с гораздо большей скоростью и отличается быстрой и легкой обратимостью реакций.
Поэтому физиологическая приспособляемость связана с более широкой «эврибионтностью», чем морфологическая приспособляемость.
Организм может жить и размножаться в весьма разнообразных условиях. При быстроте физиологической приспособляемости, и в особенности приспособляемости поведения, организм относительно легко перестраивается даже при изменении (в известных пределах) биотической обстановки. При прочих равных условиях это означает уменьшение интенсивности элиминации и увеличение численности особей данного вида и более широкое его распространение (конечно, в пределах возможности физиологической адаптации данного вида).
Жизнь большой популяции в разнообразных условиях способствует также развитию генетической гетерогенности — накоплению резервов внутривидовой изменчивости, которая, благодаря наличию весьма совершенной системы морфогенетических регуляций, имеет в основном скрытые формы. Свободная миграция и скрещивание особей различных популяций способствуют возникновению все новых комбинаций. Тому же благоприятствует и малая интенсивность элиминации индивидуальных уклонений, компенсируемых способностью к физиологической адаптации. Все это означает наличие весьма благоприятных условий для накопления большого резерва наследственных изменений и для мобилизации этого резерва при изменении внешних условий. Поэтому высокое развитие регуляторных корреляций, всегда сопровождаемое физиологической приспособляемостью организма, оказывается важным фактором, повышающим эволюционную пластичность, т. е. мобильность вида в процессе его исторических преобразований. Потенциальная способность организмов к прогрессивной эволюции достигает в этом случае наивысшего уровня.
б. Прогрессивная интеграция органических форм. Если в процессе морфологической адаптации организма к разным сторонам внешней среды создаются новые дифференцировки, то через ре-гуляторные системы и физиологические адаптации эти дифференцировки связываются в гармонически построенное и согласованно действующее целое. Создается особая пластичность организации, при которой как благоприятные наследственные изменения, так и функциональные адаптации сразу сопровождаются согласованной с ними перестройкой всей организации. Адаптации дополняются уже в процессе индивидуального развития соответственной коадаптацией органов. Таким образом, организм эво-люирует всегда как гармоническое целое и отдельные новопри-обретения не нарушают этой согласованности организации. Новые функциональные дифференцировки фиксируются, однако, как наследственное усложнение организации, лишь через стабилизирующую форму естественного отбора. Только отбор наследственных уклонений, связанных с дальнейшей дифференциацией внутренних факторов развития (определяющих развитие этих дифференцировок), может привести к такому фиксированию. Это означает, однако, дальнейшее усложнение системы корреляций.
Таким образом, в процессе эволюции наравне с прогрессивной дифференциацией идет всегда и сопутствующая ей интеграция. Происходит объединение новых дифференцировок путем их подчинения общей системе внутренних факторов индивидуального развития. Это означает усложнение системы морфогенети-ческих взаимозависимостей, а вместе с тем и прогрессивную автономизацию онтогенеза (Шмальгаузен, 1938а, б, 1942).
5. ЭВОЛЮЦИЯ ИНДИВИДУАЛЬНОЙ ПРИСПОСОБЛЯЕМОСТИ И ФОРМООБРАЗОВАНИЯ
Если результаты физиогенной и в особенности функциональной адаптации приобретают постоянное значение в известных условиях существования, то они включаются в организацию и фиксируются в результате непрерывного действия стабилизирующего отбора. Вместе с тем происходит усложнение системы морфогенетических корреляций как внутренних факторов индивидуального развития, переходящего от зависимых дифференци-ровок к установлению все более автономных самодифференцирующихся систем.
Из этого вытекает значение индивидуального приспособления как фактора, намечающего дальнейший путь эволюции. Однако индивидуальная приспособляемость и сама меняет в процессе эволюции свои формы. Способность к физиогенным адаптациям дополняется способностью к функциональным и замещается затем, по мере развития регуляторных механизмов, способностью к физиологическим адаптациям. Среди последних у позвоночных животных выдвигается приспособительное поведение как высшее выражение регуляторных функций центральной нервной системы.
А. ИНДИВИДУАЛЬНАЯ ПРИСПОСОБЛЯЕМОСТЬ КАК ЭЛЕМЕНТ ОНТОГЕНЕЗА
Индивидуальное развитие всегда является частично зависящим от факторов внешней среды. Провести определенную грань между признаками и дифференцировками, зависящими и не зависящими от внешних факторов, практически невозможно — онтогенез в этом смысле неразложим. Однако несомненно, что удельный вес таких зависимостей у различных организмов весьма различен, и если мы не можем учесть долю участия различных факторов в нормальном онтогенезе, то можно установить влияние изменений внешних факторов на формообразование организма в условиях эксперимента. Таким образом, не только вскрывается различный масштаб этой зависимости, но и различное ее выражение у различных организмов. В огромном большинстве случаев, если внешние факторы меняются в тех пределах, в которых они изменяются в естественных условиях местообитания данного организма, зависимые изменения оказываются адаптивными. Так как в природных условиях каждая особь любого вида организмов подвергается тем или иным внешним воздействиям (т. е. их изменениям по сравнению со средней нормой), то это всегда оказывает известное влияние на индивидуальное формообразование. Кроме физиогенных модификаций при этом сказывается также и значение функциональных адаптации. Каждая особь оказывается в том или ином направлении модифицированной. Таким образом, адаптивные модификации входят в состав онтогенеза любой особи — они в большей или меньшей степени определяют конкретное ее оформление, являясь как бы индивидуальной надстройкой, известным завершением ее формообразования.
Масштаб и формы этой индивидуальной приспособляемости у различных организмов весьма различны и зависят от конкретных условий их существования, а также от формы и степени активности самих организмов.
У неподвижно прикрепленных организмов приспособляемость имеет почти исключительно физиогенный характер и выражается главным образом в изменениях роста отдельных частей, в частности — в явлениях неравномерного роста, ведущего к изгибам и определенной ориентировке по отношению к внешним факторам («тропизмы»). Это наблюдается у растений, а также у сидячих животных (губки, гидроиды и др.). У растений такие модификации выражаются и в изменении формы и строения органов (световая и теневая формы листьев, строение тканей листа, величина устьиц, толщина эпидермиса, развитие опушения, ветвление, развитие механических тканей стебля и мы. др.). Способность к физиогенным адаптациям достигает именно у растений своего высшего выражения, что находится в связи с пассивными формами распространения (ветром, животными), при которых семена при их прорастании совершенно случайно попадают в те или иные условия освещения, влажности, состава почвы и т. п. Сидячих животных это не касается в такой мере, так как у них всегда имеется стадия подвижной личинки, которая прикрепляется обыкновенно лишь на более подходящих местах.
Физиологические реакции лежат в основе всех адаптации, в том числе и в основе морфогенетических адаптации. По мере усложнения организации животных они приобретают все возрастающее значение. Почти все они имеют регуляторный характер и обладают легкой обратимостью.
Это означает легкую возмещаемость потерь, связанных с физиологической деятельностью органов (физиологическая регенерация). Однако при усиленной нагрузке органа происходит не только восполнение этих потерь, но нередко и функциональная гипертрофия, до известной степени соответствующая этой усиленной нагрузке. И это также можно рассматривать как физиологическую регуляцию. Она имеет, однако, далеко идущие последствия, так как на ней основана функциональная приспособляемость органов к требуемой нагрузке. Функциональная приспособляемость органов означает возможность приспособления организма ко многим совершенно новым его положениям в окружающей среде, и притом не только в зависимости от изменений в физических факторах, но и по отношению к биотической обстановке. Быть может, еще важнее то обстоятельство, что функциональная приспособляемость простирается и на взаимоотношения между различными органами внутри организма и ведет к быстрой их взаимной приспособляемости (коадаптации) при всех индивидуальных и, тем более, исторических перестройках. Все отдельные адаптивные изменения органов дополняются сейчас же согласованным преобразованием всей организации. Организм перестраивается как гармоническое целое с тем большей легкостью, чем выше у него развита способность к функциональной адаптации и коадаптации органов.
Собственно физиологические адаптации, не проявляющиеся в постоянных изменениях структуры органов, быстро и легко обратимые, не только не теряют при этом своего значения, но, наоборот, претерпевают все далее идущие дифференцировки и достигают высшего уровня только у высших позвоночных. Основное их преимущество именно в скорости и легкой обратимости реакций. Это дает возможность преодолеть даже кратковременные изменения в факторах внешней среды (на которые организм не мог бы ответить модификацией, а если бы и ответил морфоге-нетической реакцией, то ее затрудненная обратимость привела бы к отрицательному результату после восстановления прежнего положения во внешней среде).
Поэтому развивается сложная система физиологических реакций на временные — сезонные и в особенности на кратковременные — суточные и случайные изменения в факторах внешней среды (у растений в особенности на изменения освещения, у животных — на температурные изменения).
Среди физиологических реакций у животных выделяется значение двигательных реакций, выражающихся у простейших и у низших многоклеточных в форме таксисов, а у сложных животных с центральной нервной системой — в виде сложных рефлекторных реакций и движений, основанных на инстинктах. Наконец, у высших позвоночных развивается система условных рефлексов и зачатки разумной деятельности. Приспособленность поведения достигает высшего уровня.
Мы здесь касаемся этого лишь постольку, поскольку и эти формы приспособления могут входить в состав онтогенеза. Относительно функциональных приспособлений это совершенно ясно. При определенном положении организма во внешней среде известная функциональная отшлифовка структур завершает его нормальное формообразование. У высших позвоночных — птиц и млекопитающих — это принимает иногда формы систематической тренировки («игры» и «обучение» молодых животных).
Однако и система условных рефлексов, связанных с приспособительным поведением, устанавливается также лишь в порядке тренировки и входит в состав «онтогенеза» через «обучение» молодых животных и их жизненный опыт. Эта приспособляемость поведения играет большую роль в выработке положения организма в биоценозах и установлении его взаимоотношений с другими организмами. Она означает максимальную активность организма и имеет огромное значение в борьбе за существование и в прогрессивной эволюции высших животных.
Б. АВТОНОМИЗАЦИЯ ОНТОГЕНЕЗА
В процессе эволюции происходит включение результатов индивидуальной адаптации в состав нормальной организации во всех тех случаях, когда эти адаптации получают в данных условиях существования постоянное значение положительных приобретений. Хотя эти адаптации строятся на существующей уже унаследованной норме реакций, они могут дифференцироваться далее. На базе общей нормы реакций организма возникают новые частные дифференцировки. Через стабилизирующую форму отбора происходит включение этих частных приобретений в состав «наследственных» структур. Норма реакций организма обогащается все более частными, дифференцированными формами реакций. Мы сейчас не можем сказать, насколько эволюция строилась за счет прямого отбора новых форм реагирования (ведущая форма отбора) и какова доля указанного процесса дифференцирования существующих уже реакций с последующим затем подведением наследственной базы через стабилизирующую форму естественного отбора. Дискуссия по этому поводу при настоящем уровне наших знаний была бы бесполезна.
Однако несомненен факт, что непосредственно передаются по наследству лишь половые клетки, а весь онтогенез представляет индивидуальную надстройку, осуществляемую каждый раз наново в виде цепи реакций, освобождаемых внешними факторами. Эти реакции реализуются при непрерывном взаимодействии наследственных и внешних факторов, на основе биохимических и физиологических процессов через посредство сложной системы морфогенетических взаимозависимостей.
Кроме того, мы можем утверждать, что в процессе эволюции происходит постепенное сокращение детерминирующего значения физических факторов внешней среды. Путем установления физиологических регуляций они лишаются своей специфики и, теряя свое морфогенетическое значение, низводятся на уровень условий нормального развития. Непрерывно возрастает значение внутренних факторов развития в виде системы морфогенетических взаимозависимостей. Последняя является, очевидно, некоторой надстройкой над первичными — наследственными факторами, заключающимися в организации половой клетки. В таком случае первоначальный механизм индивидуального развития должен был развертываться на основе более непосредственного взаимодействия между половыми клетками и внешними факторами.
Для эволюции реакционных и корреляционных систем в особенности характерно развитие регуляций, предохраняющих организм от нарушающих влияний различных изменений во внешних факторах. Последние «осваиваются» организмом — постоянные или закономерно повторяющиеся влияния используются организмом как источники энергии, как факторы или по меньшей мере как условия его нормального развития. С другой стороны, организм постепенно освобождается от влияний случайного и, в особенности, кратковременного характера. Это означает, что в процессе эволюции организм эмансипируется от факторов внешней среды, становясь в своих жизненных процессах и индивидуальном развитии все более автономным. Можно говорить об автономизации онтогенеза как об одном из направленных (в общем) процессов эволюции органического мира.
На низших ступенях органической жизни зависимость организма от физических факторов внешней среды была, очевидно, наиболее прямой. Через элиминацию всех уклонений от приспособленной нормы стабилизирующий отбор приводил к выработке внутренних механизмов, противодействующих неблагоприятным влияниям (регуляции) и способствующих использованию благоприятных влияний (адаптации). В процессе эволюции шло все время приобретение новых более благоприятных форм реагирования (адаптивные модификации и физиологические реакции) и установление частных, более дифференцированных реакций в порядке индивидуального приспособления.
Первоначально эти модификации (как и физиологические реакции) имели главным образом физиогенный характер, т. е. являлись вполне зависимыми изменениями. В процессе эволюции характер зависимости, однако, изменяется. Зависимые модификации дополняются регуляционными механизмами, вводящими их в определенное русло целостных, типичных изменений всей организации. Вырабатываются известные типы приспособления (адаптивные нормы), защищаемые регуляциями, органичивающими нормальную реакцию более или менее широкими пределами обычных изменений внешних факторов. Нормальная реактивность тканей устанавливается на известном уровне между нижним и верхним порогами их раздражимости. Такое развитие мы называем авторегуляторным. Одновременно с развитием системы регуляций устанавливаются, как указывалось, нижний и верхний пороги нормальной формообразовательной реакции.
Дальнейшее развитие авторегуляторной системы, определяемое элиминацией неблагоприятных уклонений, вызываемых факторами, лежащими за пределами пороговых уровней нормальной реактивности тканей, приводит к возможному их раздвиганию. В тот момент, когда оба порога реактивности раздвигаются за пределы встречающихся колебаний во внешних факторах, развитие становится автономным. Необходимые внешние факторы (ранее определявшие наступление реакции) практически всегда имеются налицо. Не только специфика, но и время наступления реакции определяются теперь внутренними факторами, а внешние факторы являются лишь условиями «нормальной» среды, в которых вообще возможно развитие организма.
Таким образом, «пусковой механизм» конкретных физиогенных адаптации, приобретших постоянное значение в жизни организма, включается внутрь самого организма. Это касается, однако, не только физиогенных адаптации. Быть может, еще в большей мере это относится к функциональным приспособлениям. Если они, возникая на базе общей способности к физиологической регенерации (функциональной гипертрофии), приобретают положительное значение в известном частном оформлении как новая дифференцировка, то они также могут быть включены в нормальную организацию через развитие внутреннего механизма этой дифференцировки.
Для нас совершенно ясно, что функциональные изменения имеют здесь руководящее значение и определяют путь дальнейшей эволюции. Роющая лапа млекопитающего может развиться только у роющего животного. Ласт может развиться только у действительно плавающего животного. Мощные челюстные мышцы могут развиться только у животного, дробящего твердую пищу или рвущего прочные ткани более крупной добычи. Приспособленные к прыжкам ноги развиваются лишь у скачущего животного. Обратный путь развития немыслим. Не может развиться ласт у наземного животного.
Однако это не исключает того, что наличие известных организационных предпосылок не только облегчает переход в иную среду, но может быть и необходимым условием, без которого такой переход был бы невозможен.
Пути, по которым идет смена факторов индивидуального развития у позвоночных животных по мере прогрессивной автономи-зации онтогенеза, уже прослежены на сравнительно-эмбриологическом материале, дополненном экспериментальными исследованиями. В этой связи я напомню работы А. А. Машковцева (1936 и др.). Оказалось, что можно установить существование вполне закономерных смен значения факторов онтогенеза в филогенетическом ряду позвоночных. В развитии вторичных почечных канальцев и мочеточника, а также в развитии легочных альвеол у низших позвоночных (у амфибий) решающую роль играет сама функция, без которой эти структуры не развиваются. При переходе к выше стоящим позвоночным формообразовательное значение функции падает и сдвигается на более поздние стадии. У аксолотля фрагментация легкого и образование альвеол идет исключительно под влиянием функции; у лягушек в дофункциональный период намечается лишь первичная фрагментация легкого. У жаб, как животных, еще более приспособленных к наземной жизни, альвеолярная структура легкого развивается еще в дофункциональный период, именно у личинки, которая у жаб легкими совершенно не дышит. В функциональный период происходит дальнейшее усложнение строения легких. Первоначальная фрагментация легкого происходит у жабы не под влиянием функции, но под влиянием гормона щитовидной железы, который определяет целый ряд морфогенетических процессов во время метаморфоза амфибий. У рептилий и у млекопитающих происходит типичное «самодифференцирование» легочных структур в течение эмбрионального периода. Функционирование легких начинается, естественно, только после рождения и ведет к дальнейшему новообразованию структур. Лишь клоачные и сумчатые млекопитающие составляют исключение в том смысле, что у них функция сохранила еще значение основного формообразовательного фактора и альвеолы легкого развиваются только под влиянием начинающегося дыхания.
Здесь ясно видно формообразовательное значение функции у филогенетически молодых органов. В дальнейшей их эволюции значение формообразовательных факторов переходит иногда к эндокринным влияниям, т. е. во всяком случае к внутренним факторам развития организма, и, наконец, зависимое развитие структур может более или менее полностью уступить место «самодифференцированию», т. е. «автономному» развитию, которое зависит в основном от внутренних факторов, заложенных в самой ткани данного зачатка (по меньшей мере после его окончательной детерминации). Картина постепенной замены внешних факторов развития внутренними, гормональными или морфогенетическими факторами и затем переноса этих внутренних факторов внутрь самого зачатка (при утрате специфичности морфогенных влияний) довольно ясна. Конечно, структура легких даже у примитивных амфибий не возникает под влиянием простого растяжения воздухом, хотя процесс фрагментации легкого у низших амфибий весьма напоминает такое пассивное растяжение стенок между развивающейся сетью кровеносных сосудов, играющих роль как бы механических препятствий. По сосудам идет затем развитие легочных трабекул. Однако у современных амфибий дело обстоит не так просто. Легкое амфибий имеет за собой длительную историю, и, несомненно, его развитие определяется уже прочной наследственной базой. Однако внешний фактор через посредство функции (и даже через механическое растяжение легочного мешка) сохранил еще значение освобождающегося раздражителя. Нужно думать, что раньше роль функции была еще выше и имела хотя бы частично и детерминирующее значение. Во всяком случае, указанный ряд форм с разным способом развития легких дает нам вполне конкретную картину замены внешних факторов развития внутренними. Эта последовательная замена в процессе эволюции одних факторов, которые послужили поводом для возникновения новых адаптивных реакций, другими — более надежными — означает стабилизацию формообразования. Такая смена факторов развития, т. е. перестройка всего механизма онтогенеза, совершается, очевидно, под направляющим влиянием стабилизирующего отбора (через элиминацию всех фенотипичеоких уклонений от приспособленной в данных условиях, т. е. адаптивно модифицированной нормы). Во многих случаях выраженного мономорфизма наиболее надежный результат дают реакции на внутренние факторы развития (ранее лишь сопутствовавшие внешним факторам как внутренние условия развития нормы). Тогда стабилизирующий отбор приводит к более или менее полной автономизации.
При наличии функциональных дифференцировок формообразовательное значение функции также, конечно, всегда дополнялось наличием ряда внутренних факторов нормального развития. И здесь устанавливались известные нормы функциональной адаптации, известный оптимум реакции и определенные пределы нормальной реактивности тканей. Менялась и реактивность тканей на различные внутренние факторы. Стабилизирующий отбор шел и здесь по пути установления наиболее надежных механизмов индивидуального развития, и если известная вполне конкретная функциональная адаптация и, в особенности, новая функциональная дифференцировка приобретала постоянное значение в данных условиях существования, то и она стабилизировалась. Функциональные факторы заменялись гормональными; гормональные факторы уступали место морфогенотическим. Локальные влияния, сопутствовавшие развитию новых функциональных дифференцировок, приобретают значение обязательных условий этого развития и при постепенном изменении реактивности самих тканей становятся детерминирующими или по меньшей мере, внутренними факторами, освобождающими автономное течение морфогенетического процесса. Все эти изменения происходили под очевидным влиянием стабилизирующей формы естественного отбора, который, следовательно, выступает в роли важнейшего агента, обусловливающего смену факторов индивидуального развития, определяющего непрерывный процесс включения индивидуальных адаптации постоянного значения в состав нормальной организации и ведущего, следовательно, к перестройке всего онтогенеза на основе прогрессивного повышения надежности нормального формообразования и устойчивости приспособленной нормы.
Подчеркивая значение стабилизации формообразования и роли стабилизирующего отбора, я, во избежание недоразумений, должен отметить, что эта стабилизация форм, в моем понимании, конечно, вовсе не исключает противоположных процессов лабили-зации. Не может быть и речи о направленном уменьшении индивидуальной лабильности и, в частности, приспособляемости форм (так же как нет и речи об уменьшении мобильности или эволюционной пластичности или о «застывании форм» в смысле Бойрлена и др. авторов). Наоборот, я подчеркиваю возрастающую пластичность органических форм и при этом обращаю внимание на возрастающее значение индивидуальной приспособляемости. Я отмечаю органическую связь процессов лабилизации и стабилизации форм. Лабилизация в общих реакциях (приобретение новых норм реакций широкого значения) сопровождается стабилизацией в частностях, т. е. автономизацией развития и включением в нормальную организацию новых дифференциро-вок. Наравне с непрерывным процессом стабилизации частных форм реагирования и, в особенности, функциональных дифференцировок, приобретающих в установившихся условиях существования постоянное значение, происходит также приобретение совершенно новых форм реагирования. На этих новых формах реагирования строится и способность к индивидуальной адаптации, в том числе — к адаптивным модификациям. Способность к более широкой адаптации означает всегда крупное приобретение, выводящее организм на новые пути прогрессивной эволюции.
6. ОНТОГЕНЕТИЧЕСКИЙ ГОМЕОСТАТ
Все биологические системы характеризуются большей или меньшей способностью к саморегуляции, т. е. гомеостазису. С помощью авторегуляции поддерживается само существование каждой данной системы, ее состав и структура с ее характерными внутренними связями и закономерными преобразованиями всей системы в пространстве и времени. Гомеостатическими системами являются, конечно, прежде всего отдельная особь каждого вида организмов, затем популяция как система особей одного вида, характеризующаяся своим составом и структурой с особыми взаимосвязями ее элементов, и, наконец, биоценоз, обладающий также определенным составом и структурой со своими подчас очень сложными взаимозависимостями.
Устойчивость онтогенеза, ведущего к сформированию нормальной организации, определяется в основном сложной системой взаимозависимостей, которые имеют более или менее регу-лятарный характер. Сущность регуляторных механизмов еще недостаточно изучена.
Под регулированием вообще понимают действия, направленные на поддержание системы в требуемом состоянии. В частности, оно включает поддержание заданного необходимого или подходящего движения, его направления и скорости (течения, передачи энергии, химических реакций и т. п.), поддержание заданных соотношений в этом движении и поддержание подвижного равновесия. Все эти явления могут быть взаимосвязаны. Способы регулирования можно разграничить по устройству соответствующих механизмов и применяемым при этом средствам.
В технике различают регулирующие механизмы разной сложности: 1) простое регулирование по заданной программе, 2) прямое — с учетом факторов, вызывающих уклонения от программы, и 3) регулирование по замкнутому циклу с обратной связью, т.е. с проверкой результатов регулирования по сравнению с заданной программой и внесением соответствующих поправок. Последняя, высшая форма регулирования включает и первые два механизма.
В онтогенетическом процессе можно установить наличие тех же принципов регулирования: 1) развитие по программе, заданной наследственным материалом, т. е. внутренними факторами развития; 2) развитие соответственно положению во внешней среде, т. е. зависимость от внешней среды и 3) регуляция в собственном смысле, т. е. выправление уклонений и восстановление «нормальных» соотношений при их нарушениях. Последнее возможно лишь через посредство замкнутого цикла зависимостей, т. е. при наличии обратной связи между развивающейся частью и наследственной основой норм реагирования.
Регулирование с обратной связью может быть сравнительно простым в пределах одной частной системы, но может достигать и большой сложности во взаимодействующих системах со взаимным контролем результатов. Формы взаимодействия могут быть также различными. Частные системы могут в своем взаимодействии оказывать стимулирующее влияние друг на друга. Это ведет к согласованному последовательному развитию этих частей (положительная обратная связь). Возможно и такое соотношение, когда одна часть оказывает на др*угую стимулирующее воздействие, а вторая часть — на первую, задерживающее (отрицательная обратная связь). Это ведет к установлению стационарного состояния.
Цикл регуляции с обратной связью может быть соединен с другим подобным циклом, контролирующим и выправляющим работу первого. Такое регулирующее устройство называется самонастраивающимся.
Большое значение для регуляции соотношений имеет также возможность противоположного воздействия на один и тот же процесс — возбуждение и торможение. В биологических системах противоположные воздействия передаются обычно по разным каналам (по различным нервам или от разных органов внутренней секреции, или, (во всяком случае, разными физическими или химическими средствами). В сформированном зачатке и в дифференцированном организме в целом наличие таких взаимозависимостей ведет к поддержанию известных соотношений.
Программное регулирование детерминировано унаследованной нормой реагирования в определенных условиях. Оно осуществляется в пределах этой нормы в некоторых модификациях, определяемых обычными изменениями во внешних факторах (прямое регулирование). Однако при кратковременных или необычных уклонениях во внешних факторах и даже во внутренних факторах развития (т. е. в самой программе) возможно все же нормальное формообразование вследствие наличия защитных механизмов в виде пороговых уровней нормальной реактивности тканей и запаса реагирующего материала. Реакция наступает лишь в том случае, если специфический раздражитель достигает известного минимального уровня интенсивности и, раз начавшись, протекает до конца за счет внутренних запасов реагирующего материала. Нормальный результат обеспечивается и при известных нарушениях соотношений вследствие наличия избыточного количества реагирующих веществ. За счет последних происходит компенсация возможных дефектов. Примером такого действия физиологических механизмов может служить поддержание активной реакции крови на постоянном уровне за счет ее забуференности. В онтогенетических системах поддержание нормального фенотипа при обычных мутациях осуществляется за счет доминирования нормы. В этом случае развитие нормы обеспечивается наличием двойной дозы данного гена в гомозиготе и блоком полигенов или комплексом модификаторов в гетерозиготе. При изменении одного гена его аллеломорф или сам по себе или в соединении с комплексом модификаторов оказывается достаточным для осуществления нормы. Весьма надежную защиту дают полигенные системы, обладающие «запасом» в виде целой серии генов со сходным действием. В морфогенети-ческих системах обычно даже значительно уменьшенная доза индуктора вызывает полноценную реакцию. Точно так же материал, способный к специфической реакции (например, линзооб-разование), всегда намного превышает то количество, которое используется в нормальном формообразовании.
Компенсация достигается нередко и избыточной организацией самой формообразовательной системы. Обычно детерминация и формообразование определяются не одним каким-либо взаимодействием (индукцией), а более сложными взаимовлияниями. В механике развития некоторые такие явления известны под названием «двойного» обеспечения.
Таким образом наличие двойной дозы гена, полигенной обусловленности, наличие избытка индуцирующего и реагирующего материала и, наконец, множественной обусловленности формообразовательных реакций — все это оказывается средствами, защищающими нормальное течение онтогенеза от возможных нарушений под влиянием изменений во внешних или внутренних факторах. Такие защитные механизмы, основанные на компенсации за счет имеющихся запасов, можно также называть регуляторными механизмами в широком смысле, так как они делают программное регулирование более надежным.
Более совершенная регуляция возможна за счет соответствующей организации формообразовательной системы, которая автоматически перестраивается при изменении какой-либо ее части.
Элементарный регуляторный механизм осуществляется в простых зачатках с полярной организацией, характеризующейся количественными различиями в определенном направлении, т. е. наличием известного «градиента» в распределении веществ и в интенсивности метаболизма.
Сложные регуляторные механизмы построены в организмах всегда по типу замкнутого цикла с обратной связью. Они наблюдаются как в физиологических регуляциях, так и в морфологических процессах.
Примеры физиологических и морфологических регуляций: мышечное сокращение определяется возбуждением двигательных нервных центров и контролируется рецепторами в самой мышце, от которой возбуждение передается на чувствующий, а оттуда вновь на двигательные нервные центры; морфогенетические регуляции — корневая система обеспечивает минеральное питание и через это определяет развитие листьев и размеры ассимилирующей их поверхности, а ассимиляция в листьях определяет рост корневой системы; глазной бокал вызывает образование хрусталика, хрусталик — дифференцировку ретины.
Эти примеры морфогенетических регуляций показывают взаимную стимуляцию зависимых процессов, ведущую к прогрессивному развитию всей системы. Возможно, однако, и взаимодействие с обратным направлением. Например, функция семенников стимулирует у петухов развитие гребня, а последний тормозит дальнейшее развитие семенников. Если удалить гребень, усилится рост желез. Если удалить железы — редуцируется гребень (М. Завадовский,1922).
К сожалению, эти системы взаимного контроля и регуляции соотношений в развивающемся организме еще недостаточно изучены. Однако не подлежит сомнению, что явления авторегуляции характерны для всех жизненных проявлений вообще, а в частности имеют большое значение и в индивидуальном развитии организмов. В процессе эволюции, с усложнением организации, меня ются, усложняются и совершенствуются и регуляторные механизмы. Вместе с защитными механизмами (характера компенсации) они обеспечивают развитие нормы при наличии случайных уклонений во внешних и внутренних факторах. Обычные, небольшие колебания в факторах среды, а также малые мутации не проявляются как-либо на формообразовании (во всяком случае в существенных признаках). Индивидуальное развитие нормы приобретает максимальную устойчивость, но вместе с тем и известную приспособляемость к закономерным изменениям в факторах внешней
среды.
Эволюция организмов определяется их меняющимся положением во внешней среде, выражающимся сложными взаимозависимостями, которые были Ч. Дарвином названы борьбой за существование. Обязательными проявлениями последней являются известная степень индивидуального соревнования и элиминация более или менее значительного числа особей, как следствие высокой потенциальной размножаемости организмов. В процессе прогрессивной эволюции наибольшее значение приобретает возрастающая активность организма, так как именно активная борьба с вредными влияниями и активная конкуренция связаны с наиболее резко выраженной избирательной элиминацией. Преобладание пассивных форм соревнования связано у низших организмов с весьм:а значительной общей элиминацией случайного характера. Увеличение активности высших животных сопровождается поэтому возрастанием эффективности естественного отбора.
В направляющем действии естественного отбора мы различаем прежде всего его ведущую роль, основанную на селекционном преимуществе известных уклонений от нормы перед установившейся нормой, теряющей свою приспособленность в новых* условиях существования. Она ведет к установлению новых норм реакций, реализующихся в новых адаптациях организма и в новых формах индивидуального приспособления. С другой стороны, мы отмечаем особо стабилизирующую роль естественного отбора, основанную на селекционном преимуществе приспособленной уже нормы перед всеми от нее уклонениями. Так как отбор идет по фенотипам, то элиминация уклонений охватывает как мутации с заметным выражением, так и модификации, оказавшиеся в данных условиях неблагоприятными. Элиминация результатов «ошибочных», «преждевременных» реакций на случайные и кратковременные уклонения в факторах внешней среды ведет к отбору особей с более устойчивым формообразованием, обладающих, между прочим, ограниченной реактивностью тканей и внутренними регуляторными механизмами, противодействующими вредным влияниям факторов внешней среды. Такими механизмами являются: 1) диплоидность, практически гарантирующая наличие двойного комплекта нормальных аллеломорфов в каждой особи и нейтрализующая до известной степени влияние отдельных гетерозиготных мутаций, 2) доминантность нормы, гарантирующая устойчивость нормального формообразования при наличии мутаций, 3) сбалансированность генетической системы, устанавливающейся через отбор наиболее жизненных комбинаций, 4) устойчивость морфогенных влияний п реактивности тканей, специфика которых определяется не отдельными генами, а всем генотипом в целом, 5) построение морфогенетических взаимозависимостей (в виде градиентов, индукционных систем и других морфогенных и гормональных влияний), допускающих существование значительных сдвигов во времени и скорости поступления и в изменениях концентрации морфогенных веществ, а также сдвиги во времени созревания и в уровне нормальной реактивности тканей, 6) защита нормальной реактивности тканей путем установления широкого диапазона возможных изменений в интенспвностях внешних факторов или в концентрации морфогенных веществ (т. е. в интен-сивностях внутренних факторов) без нарушения нормального формообразования,— это означает возможно широкое раздвигание нижнего и верхнего порогов нормальной реактивности тканей, и 7) сложность системы морфогенетических корреляций, не допускающая разрушения отдельных ее звеньев без летальных последствий и приобретающая все более ярко выраженный регуляторный характер.
Все эти механизмы развиваются, очевидно, под направляющим влиянием стабилизирующей формы естественного отбора. Их наличие и уровень развития имеют огромное значение в процессе дальнейшей прогрессивной эволюции организмов.
Эволюция форм реагирования ведет от начальных форм онтогенеза, зависящего от внешних факторов, к авторегуляторному развитию и, наконец, ко все большей степени автономности.
Развитие регуляторных систем и прогрессивная автономизация развития означают полную перестройку онтогенеза со всеми его факторами. Основное значение имеет при этом прогрессивное усложнение системы морфогенетических корреляций, создаваемых за счет элементарных выражений плейотропизма и постепенно приобретающих все более выраженный регуляторный характер. При этом достигается максимальная эволюционная пластичность организма, как за счет большого скрытого резерва изменчивости и связанной с нею мобильности, так и за счет индивидуальной приспособляемости и возможности быстрого фиксирования ее важнейших результатов. Это обеспечивает максимальные темпы эволюции высших животных с их большой индивидуальной приспособляемостью и автономным развитием, при высоко развитой системе регуляций.
Указанная перестройка онтогенеза идет под направляющим влиянием стабилизирующей формы естественного отбора, получающей по мере эволюции органического мира все возрастающее значение.