
Проблемы Эволюции |
В тени дарвинизма. Альтернативные теории эволюции в ХХ веке. СПб: Ясный день, 2003. С. 158-175.
Попов И. Ю.
Загадка теории эволюции - происхождение рукокрылых
В тени дарвинизма. Альтернативные теории эволюции в ХХ веке. СПб: Ясный день, 2003. С. 158-175.
The mystery of evolutionary theory – the origin of bats
Small animals similar to modern insectivores or lower primates are considered to be the starting point of the evolution of mammals. The transitive forms between such “non-specialized” animals and Chiroptera are not known, and it is difficult to imagine them. The idea suggests itself, that their ancestors had underdeveloped wings. Those wings however, which could not be used for flight, might lower the fitness of the animal carrying them. This means, that to characterize the process of the origin of Chiroptera it is necessary either to explain how these unfit organisms could arise and evolve, or to substantiate the possibility of the absence of the transitive forms, or to prove the adaptive value of the underdeveloped wings and other features of the hypothetical ancestors.
“The mystery” demonstrates clearly the difficulties of evolutionary theory and could aid in understanding the motives of scientific anti-Darwinism.
It is possible to characterize the environment, in which the evolution of the Chiroptera ancestors took place. At the present time the rain forests of Southern Asia, i. e. the forests with the low-density highest arboreal tier, exemplify such an environment. In these forests various gliding animals have evolved several times, and maybe flapping flight originated in similar conditions. The origin of bats (one of the two groups of Chiroptera, which probably had different non-flying ancestors) was related to having a cave habitat. The capacity for echolocation indicates this fact. Various modern animals have several separate features of the hypothetical chiropteran ancestors. Skin membranes of every possible degree of development, elongated fingers, and hypertrophy of the forelimbs occur among arboreal mammals. In most cases these features could be used during jumps or gliding, and seem to be indirect evidence for the possibility of the existence of forms intermediate between Chiroptera and “non-specialized” mammals. The details of the evolution of the entire complex of chiropteran features remain unclear. An attempt to characterize them from the viewpoint of different evolutionary concepts, “the mystery” relates to the problem of the mechanisms of “tendencies” or “regularities” of variation. This problem is stated especially clearly by the concepts of orthogenesis. Moreover, the analysis of “regularities” using John Willisґ hypothesis on “Age and Area”, and biogeographic research, seems to be promising ways of solving “the mystery”. It turns out that biogeographic analysis can throw light on the origin of gliding animals. The comparative methods have been unable to solve the problem of Chiropteran origins.
Исходной формой для эволюционного развития плацентарных млекопитающих считаются мелкие четвероногие животные, похожие на современных насекомоядных или низших приматов. Переходные формы между ними и рукокрылым не только не известны, но даже не ясно, какими они могли бы быть. Напрашивается мысль о том, что они имели не вполне развитые крылья. Однако такие крылья, которые еще не могут использоваться для полёта, казалось бы, должны только мешать их обладателям и снизить их конкурентоспособность. Значит, чтобы охарактеризовать процесс происхождения рукокрылых нужно или объяснить, почему в эволюции могли появиться и развиваться неприспособленные организмы, или обосновать возможность отсутствия переходных форм, или всё же показать функциональное значение недоразвитых крыльев и других особенностей возможных предков рукокрылых.
Происхождению рукокрылых посвящена весьма обширная литература, но она касается в основном филогенетического аспекта этой проблемы, то есть выяснению родственных связей разных групп рукокрылых между собой и с их возможными ближайшими родственниками - приматами, насекомоядными и шерстокрылами (Pettigrew, 1986; Wible & Novacek, 1988; Ammerman & Hillis, 1992; Simmons, 1994; Teeling et al., 2000 и др.). Вопрос о механизмах эволюции, которые привели к появлению рукокрылых, обычно обходится молчанием. Нередко высказывалось мнение о том, что любые соображения на этот счёт - чистая спекуляция. По нашему мнению, это не совсем так. Кроме научных спекуляций, здесь можно привлечь некоторые косвенные данные, то есть данные о происхождении других летающих животных и о морфологических особенностях млекопитающих, которые могли бы продемонстрировать стадии эволюционного развития ближайших предков рукокрылых.
Какими могли бы быть переходные формы от первых плацентарных к рукокрылым и были ли они вообще? В каких условиях они могли существовать? Как объясняют их появление с разные эволюционные концепции? Этим вопросам посвящена настоящая статья. Они хорошо демонстрируют сложности современной эволюционной биологии и помогают понять мотивы научного антидарвинизма.
Происхождение полёта у позвоночных
Вопрос о происхождении любых летающих позвоночных всегда был очень трудным для теории эволюции. Поэтому его анализ нередко был связан с "ересями" в эволюционизме или даже с антиэволюционизмом. "Искать переходные звенья бесполезно, потому что их и не было – первая птица вылупилась из яйца рептилии" - это известное высказывание немецкого палеонтолога Отто Шиндевольфа (O. H. Schindewolf, 1896-1971) является основной идеей группы сальтационистских концепций, претендующих на замену современного дарвинизма. Это мнение можно считать преувеличением, потому что между птицами и рептилиями есть хоть какие-то переходные формы. Они рассматриваются как "боковые ветви" эволюции, но, по крайне мере, демонстрируют некоторые стадии процесса происхождения птиц. Но в случае рукокрылых действительно создаётся впечатление внезапного возникновения группы летающих животных без всяких переходных форм. Самая древняя из ископаемых летучих мышей мало отличается от современных, хотя жила около 60 миллионов лет назад.
Тем не менее предложено несколько гипотез происхождения полёта среди позвоночных, в которых анализируются именно переходные формы от нелетающих животных к летающим. Гипотезы можно разделить на две группы: в одних полёт рассматривается как развитие прыжков вверх, а в других - прыжков вниз. Первая группа гипотез относится в первую очередь к птицам. Некоторые авторы считают, что ближайшие предки птиц могли быстро бегать и высоко прыгать. В ходе эволюции они приобретали способность прыгать всё выше и выше, параллельно обзавелись перьями и, в конце концов, полетели. Считается, что на ранних стадиях данного эволюционного процесса перья могли служить сачками для ловли насекомых или использоваться для управления прыжком (Padian, 1985; Norberg, 1990). Подобная версия происхождения полёта птиц отчасти подтверждается тем, что известны динозавры, которые сходны по анатомии с птицами, имели перья и, по всей видимости, могли быстро бегать и высоко прыгать (см. Wellnhofer, 2002). Многие современные птицы также способны к такому способу передвижения, и в их анатомическом строении нетрудно усмотреть указание на "бегающих" предков. В анатомии рукокрылых таких указаний нет. Они не только не способны к беганию, но и вообще с трудом передвигаются по земле (за исключением вампиров). Тем не менее и в отношении рукокрылых также высказана гипотеза происхождения полёта "снизу вверх". Одним специалистом по рукокрылым – К. К. Панютиным (Панютин, 1980) - высказано предположение, что предки летучих мышей были "прыгающими" и напоминали рыб - илистых прыгунов (сем. Periophtalmidae). Илистые прыгуны обладают способностью ловко передвигаться по суше и даже лазать по деревьям. Они двигаются, изгибая тело и опираясь на грудные плавники. Панютин считает, что предки летучих мышей передвигались таким же образом и тоже жили около воды в условиях, подобных мангровым зарослям.<![if !supportFootnotes]>[1]<![endif]>
Согласно гипотезам другой группы, предки летающих животных – птиц, рукокрылых, птерозавров - жили на деревьях и были способны к парашютирующим прыжкам. Крылья развивались сначала как органы, использующиеся для планирования, но со временем они настолько усовершенствовались, что стало возможным использование их и для активного машущего полёта. В современной науке предпринимались попытки развить эту идею с использованием аэродинамических расчётов. Результаты этих расчётов противоречивы. Одни специалисты говорят, что между планирующим и настоящим полётом лежит непреодолимая пропасть, так как планирующий полёт – это падение вниз, которое никак не может преобразоваться в ходе эволюции в движение наверх. Другие утверждают, что здесь нет ничего невозможного (см. Norberg, 1990). От гипотезы "древесного" происхождения полёта не вполне отказались даже при анализе происхождения птиц, несмотря на открытия "бегающих" переходных форм. Можно попытаться развить её в отношении рукокрылых. Возможно, нам удастся как-то продвинуться в данном вопросе, если мы обратимся к анализу современных планирующих животных.
Планирующие позвоночные
В современной фауне известно несколько видов млекопитающих, рептилий и земноводных, способных к планирующему полёту. Самой обширной группой планирующих животных являются белки-летяги. Это семейство грызунов (Pteromyidae), включающее около 50-и видов. Летяги внешне похожи на белок, но отличаются тем, что имеют перепонки между выступающими частями тела. Похожи на белок и на белок-летяг африканские грызуны – шипохвосты или чешуехвостые белки (сем. Anomaluridae). Они имеют интересную особенность – шипы или чешуи на нижней части хвоста, которые дают возможность их обладателям прочно удержаться на голом стволе дерева. Как и в случае белок, среди шипохвостов есть "нормальные" (р. Zenkerella) и "летающие" (р. Anomalurus). В семейство шипохвостов включают ещё одну небольшую группу планирующих животных – "планирующие мыши" (p. Idiurus)(несмотря на то что у них хвосты вполне нормальные). Кроме летающих белок и мышей, среди млекопитающих есть более специализированные планирующие животные – шерстокрылы. Их включают в отдельный отряд (Dermoptera). Как это часто бывает, у этих плацентарных млекопитающих есть аналоги среди сумчатых. В Австралии и на островах Индонезии обитают летающие кускусы или поссумы (сем. Petauridae). Три группы сумчатых развили эти способности (см. Фауна мира, 1990).
Таким образом, планирующие животные произошли независимо в нескольких группах млекопитающих, в 7-8 по крайней мере. Кроме млекопитающих, способность к планированию имеют некоторые рептилии и амфибии. Известны "летающие" змеи (род украшенные древесные змеи, Chrysopelea), "летающие" лягушки (род Rhacophorus) и "летающие" ящерицы (род Draco). В общей сложности, планирующих рептилий и амфибий в современной фауне тоже несколько десятков видов.
Все эти животные распространены неравномерно по областям Земли. Больше всего их в экваториальных или тропических лесах. Это совсем неудивительно. Джунгли представляются самым подходящим местом для древесных животных. Удивительно то, что они живут не на всех континентах. Максимальное их количество – в Юго-Восточной Азии и Северной Австралии. На крайнем юго-востоке Азии пересекаются ареалы почти всех родов белок-летяг, а также шерстокрылов, "летающих" ящериц, "летающих" змей и "летающих" лягушек. В Африке планирующих животных гораздо меньше – несколько видов шипохвостов и планирующих мышей. А в экваториальных лесах Америки планирующих животных вовсе нет. Зато там очень много животных с цепкими хвостами.
Почему же в Азии способность к планирующему полёту развивается у всех позвоночных, у кого только это возможно, а в Америке – ни у одного? Казалось бы, условия в экваториальных лесах разных континентов очень сходны, и поэтому жизненные формы их обитателей тоже должны быть сходны. Но тем не менее обнаружились существенные различия. К этому примеру иногда обращаются, чтобы проиллюстрировать "географический стиль", непонятный в привычных терминах приспособления (Чайковский, 1990). Но, как показывает опыт истории науки, в большинстве случаев, случайное событие по мере исследования всё больше кажется не очень случайным или даже жёстко предетерминированным или, по крайней мере, связанным с рядом других.
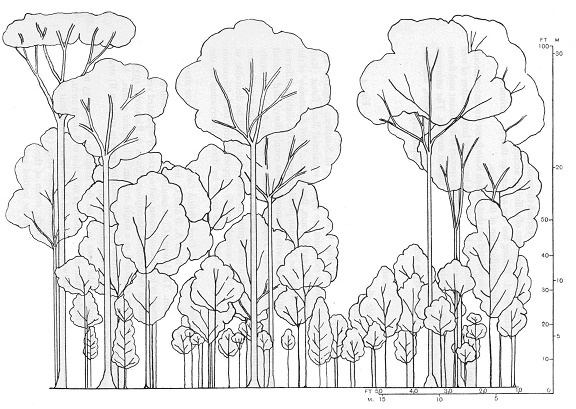
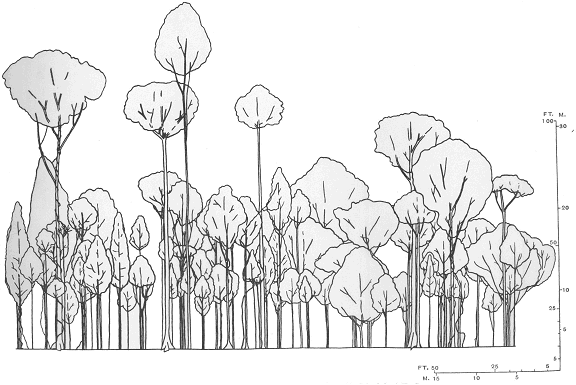
Несмотря на то что условия жизни в экваториальных лесах действительно сходны - буйная густая растительность, которая заполняет несколькими ярусами большое пространство – они всё же не одинаковы. Здесь стоит обратить внимание на различия в характере ярусности (Richards, 1958). В экваториальных лесах выявляются три древесных яруса, и различия в их развитии чётко соответствуют наличию или отсутствию планирующих животных.
В лесах Южной Америки довольно много деревьев в самом верхнем ярусе, и "окна" в нём часто бывают заполненными деревьями второго яруса. Оба эти яруса образуют густой сомкнутый полог значительной толщины. Ясно, что в таком лесу умение прыгать на несколько десятков метров останется нереализованным. Зато умение цепляться хвостом за ветки очень может пригодиться. В азиатских экваториальных лесах также есть сомкнутый полог, но его образуют нижние ярусы, а верхний сильно разрежен. Такому типу ярусности вполне соответствует то обстоятельство, что многие животные, населяющим кроны самых высоких деревьев, имеют способность к планированию. В Африке сомкнутость крон наблюдается в нижних ярусах. Расстояние между деревьями в верхних ярусах бывают сравнительно большими, и вокруг стволов остаётся много свободного пространства. Как раз там и живут шипохвосты (рис. 1).
Таким образом, анализ распространения и условий обитания планирующих животных даёт основания охарактеризовать условия, в которых все, кто только может, развивает в себе способность к полёту. Этими условиями являются экваториальные леса с разреженным верхним древесным ярусом. Попробуем соотнести эти соображения с тем, что нам известно о рукокрылых.
а
б
в
Рис. 1. Схемы ярусности экваториальных лесов Америки (а) Азии (б) и Африки (в) по П. Ричардсу (Richards, 1958).
Возможные предки рукокрылых и условия, в которых они могли появиться
Рукокрылых разделяют на две хорошо группы – крыланы и летучие мыши. Они сильно различаются, и неоднократно высказывалось предположение об их независимом происхождении от разных нелетающих предков. Крыланы близки к приматам, летучие мыши - к насекомоядным. Об этом свидетельствует сходство в строении скелета, а также нервной и репродуктивной систем. Различия есть и по некоторым хорошо заметным внешним и поведенческим особенностям. Летучие мыши питаются, в большинстве случаев, насекомыми, и имеют хорошо развитую систему эхолокации. Крыланы растительноядны, имеют лучше развитое зрение, а способность к эхолокации встречается среди них в качестве исключения, и развита намного хуже, чем у летучих мышей. У крыланов есть хорошо развитый коготь на крыле, у летучих мышей его нет (см. Hill & Smith, 1984). Некоторые исследователи именно на основании морфологических различий между крыланами и летучими мышами приходили к выводу о дифилетичном происхождении рукокрылых. Если это верно, то нужно искать уже не одного, а двух нелетающих предков рукокрылых (Pettigrew, 1986) и соответственно два ряда переходных форм. Хотя в то же время некоторые морфологические данные – например, особенности строения костей черепа - свидетельствуют о монофилетичном происхождении группы рукокрылых (Wible & Novacek, 1988).
Современные методы анализа биохимической изменчивости, (как это обычно происходит) внесли серьёзные коррективы в филогенетические схемы развития рукокрылых. В свете этих данных морфологические особенности рассматриваются как нечто вторичное, в то время как данные о молекулярной эволюции – как отображение подлинной истории развития группы. Сравнительные исследования ДНК, РНК и различных белков рукокрылых проведены в большом объёме (обзор: Pettigrew, 1999), и обычно они свидетельствуют в пользу монофилии. Но по этому поводу остаются сомнения. В последних работах этого направления изображается более сложная картина истории развития рукокрылых: летучие мыши - полифилетичная группа, часть которой близка к крыланам. Но при использовании различных маркёров разные семейства летучих мышей оказываются родственными крыланам. При обобщении этих данных был сделан вывод о том, что "все 4 или 5 сценариев полифилии летучих мышей не могут быть достоверны, но возможно, что все они не верны!" (Pettigrew, 1999. P. 3).
Сведения о ближайших родственниках рукокрылых не менее противоречивы. Их искали среди приматов, насекомоядных и шерстокрылов, объединяя все эти группы в особый надотряд Archonta. Но о положении рукокрылых в этом надотряде высказаны и подкреплены фактами почти все мыслимые точки зрения: все рукокрылые наиболее близки к насекомоядным (Kovtun, 1987); по крайней мере, часть рукокрылых – это «летающие приматы» (Pettigrew, 1986); рукокрылые не имеют особой близости ни к одному из указанных отрядов (Wible & Novacek, 1988) и др.
Таким образом, об истории развития группы нет даже какой-то наиболее распространённой точки зрения, и даже если остановиться на какой-нибудь версии, то она абсолютно ничего не говорит о возможных предках рукокрылых. Сравнительный метод в приложении к этой группе зашёл в тупик, и это даёт основание усомниться в результативности его теоретической основы. В цитированных выше исследованиях использованы, в основном, два подхода. Первый заключается в выявления сходных "значимых" признаков между группами. К примеру, определённый тип связи сетчатки со средним мозгом есть только у крыланов и у приматов. Отсюда был сделан вывод о том, что они тесно связаны между собой по происхождению (Pettigrew, 1986). В каждом подобном случае высказывается возражение: любые морфологические особенности могут возникнуть конвергентно, и поэтому едва ли их можно использовать таким образом. Другой, более современный подход заключается в расположение групп по принципу сходства-различия с учётом множества признаков (желательно биохимических), то есть чем больше сходства между группами, тем ближе они на филогенетической схеме. Эта процедура основана на допущении постоянства скоростей эволюции, положенных в основу анализа признаков, что весьма сомнительно. Как и первый подход, он не учитывает, или, по крайней мере, имеет тенденцию скрывать конвергенции (см. Moore & Willmer, 1997).
Несмотря на слабые места в теоретическом обосновании, подобные филогенетические схемы используются, поскольку трудно предложить альтернативы. Если всё же попытаться это сделать, то, возможно, продуктивным будет использование гипотезы Дж. Виллиса (J. Willis, 1868-1958) о "возрасте-ареале" или "пространства-времени". Её суть состоит в следующем. Первые представители группы высокого таксономического ранга появляются очень быстро, а потом они медленно расселяются. По мере расселения они разделяются на всевозможные подгруппы. В естественных условиях расселение идёт очень медленно, поскольку новые формы должны как-то прижиться на уже занятой территории. Отсюда следует, что группы, имеющие обширный ареал – древние, и наоборот, если ареал группы невелик, то значит, она является относительно молодой (Willis, 1922, 1940)<![if !supportFootnotes]>[2]<![endif]>.
Эта гипотеза хорошо согласуется с изложенными выше соображениями об условиях появления планирующего полёта. Распространение одной группы рукокрылых – крыланов - демонстрирует закономерность, сходную с той, которую мы показали для планирующих животных. Несмотря на то, что препятствий для расселения летающих животных, как будто бы, практически нет, они так же неравномерно распределены по континентам. Больше всего крыланов в Азии, меньше в Африке, в Америке нет совсем. Случайно ли это совпадение? На данной стадии исследования ещё можно считать его случайным, поскольку данных об истории развития и расселения крыланов ещё недостаточно. Но и существование «закономерности» здесь тоже можно предположить. Возможно, что различия ярусности лесов разных континентов были сходными и в далёком прошлом. Предки крыланов также населяли верхний разреженный ярус тропических лесов и были способны к планирующему полёту. Они, вероятно, появились значительно позже летучих мышей, поскольку их ареал относительно невелик.
О летучих мышах сведения менее определённы. Они распространены повсеместно. Можно только сказать, что на какой-то ранней стадии их эволюция была связана с пещерами. Об этом можно судить по тому, что все они имеют хорошо развитую систему эхолокации. Более того, есть указание на существование эхолокации у самых древних известных рукокрылых (Novacek, 1985). Эта особенность кажется явной адаптацией к ночному образу жизни. Но в то же время известно, что многие, если не большинство ночных животных легко без неё обходятся. Кроме того, некоторые авторы утверждают, что адаптивная ценность эхолокации сомнительна: соответствующая система органов увеличивает вес, ограничивает размеры, и поэтому крыланы, к примеру, её утратили (см. Speakman & Racey, 1991; Rayner, 1991; Teeling et al., 2000). Система эхолокации максимально развивается в том случае, если животное обитает в абсолютной темноте (где нет даже лунного света) и перемещается в трёхмерном пространстве - летает или плавает. Эхолокация как необходимый способ ориентации в пространстве есть у китообразных, у летучих мышей и у некоторых птиц, гнездящихся в пещерах. Способность к эхолокации может появиться в результате тренировки у других животных и у человека, но летучие мыши и китообразные в этом отношении стоят на недосягаемой высоте (см. Константинов, 1980).
Возможно, в таком случае, изучение животных пещер может как-то прояснить вопрос. Пещерные биотопы, конечно, являются редкими, но всё же это - неотъемлемая часть биосферы, а не случайность в геологической истории. Многие пещеры населены причудливыми рыбами, ракообразными и насекомыми. Пещерные жители имеют ряд специфических особенностей: редукция зрения, бледная окраска, сильное развитие органов осязания и т.д. Исследования фауны пещер иногда называют аномалией в эволюционной биологии. Как и любая аномалия она рассматривается или как курьёз, недостойный серьёзного внимания, или наоборот, как окно в изучение тончайших механизмов эволюционного процесса. Дарвин (C. Darwin, 1809-1882) считал этих животных реликтами древней фауны, которые сохранились в условиях низкой конкуренции. Но впоследствии это не подтвердилось: было установлено относительно недавнее происхождение многих "заселённых" пещер. Результаты первых исследований пещерной фауны приводились для обоснования ламаркизма. Появление своеобразных пещерных жителей объяснялось упражнением и неупражнением органов. Впоследствии от ламаркизма постепенно отказались. Позже было высказана гипотеза о том, что в специфических условиях пещер выявляются некие скрытые возможности эволюции, которые не могут развиться в должной мере в нормальных условиях. Такими направлениями были тенденция к утрате зрения, к удлинению плавников у рыб (см. Vandel, 1964; Bocking, 1988). Эти "тенденции" привели к появлению у животных признаков, аномальных по сравнению с особенностями их ближайших родственников. Вполне возможно, что такой же аномалией является и строение летучих мышей, т. е. к нему привели тенденции к гипертрофированному развитию передних конечностей, перепонок и т. п.
Таким образом, об условиях, в которых произошли рукокрылые, можно судить достаточно определённо. Скорее всего, это лес с определённым типом ярусности и пещеры. Но как быть с переходными формами? Если представить себе летучую мышь с недоразвитыми, зачаточными крыльями, то она может показаться нежизнеспособной. Может ли зачаточная перепонка оказаться полезной или невредной для организма? Для того чтобы ответить на этот вопрос обратимся к современной фауне.
У множества млекопитающих есть перепонка между пальцами. Обычно это связано с водным образом жизни, но не обязательно. Некоторые водные животные их не имеют (водяные землеройки, сем. Potamogalidae, например) и наоборот, перепонка есть у животных, которым плавать вовсе необязательно (например, у гиббонов Symphalagus syndactylus, сем. Hylobatidae). Зачаточная перепонка между конечностями есть у лемуров индри (род Propithecus). Как она используется – не совсем ясно, возможно, как тормозящий парашют при прыжках. Дальнейшие стадии развития кожных складок – между конечностями, а затем, между конечностями, шеей и хвостом - демонстрируют разные виды белок-летяг, шипохвостов, кускусов, поссумов. Максимального развития эта особенность достигает у шерстокрылов. У них кожная перепонка охватывает шею, конечности, пальцы и хвост. Для "превращения" в рукокрылых им не хватает непропорционального увеличения передних конечностей и удлинения пальцев. Гипертрофия передних конечностей и удлинённые пальцы есть у других животных – у обезьян, например, или у водных млекопитающих.
Таким образом, все признаки возможных предков рукокрылых по отдельности современная фауна демонстрирует, и уже поэтому принципиальных препятствий для построения ряда переходных форм нет. Среди млекопитающих, которые ведут древесный образ жизни, можно видеть перепонки любой степени развития. В большинстве случаев они находят какое-то применение при прыжках или планирующем полёте. Были высказаны и другие соображения по поводу использования кожных перепонок - сачки для ловли насекомых, ласты, теплообменники или "солнечные батареи" (Crompton, 1968; Панютин, 1980; Ковтун, 1988, 1990). Животные с подобными "инструментами" выглядели бы совершенно нелепо, хотя надо признать, что представить чисто научное опровержение этих гипотез трудно. Можно только сказать, что планирующий или парашютирующий вариант выглядел бы более "естественно", потому что он напоминает некоторых современных млекопитающих.
К этим соображениям о переходных формах можно добавить уточнения, если сопоставить их с некоторыми явлениями, описанными в палеонтологии. Даже в отношении хорошо изученных в палеонтологическом отношении групп переходные формы вызывают множество споров. Когда строят родословные деревья, все известные из палеонтологической летописи формы обычно представляют в виде боковых ветвей где-то сбоку от основного ствола эволюционного развития. Трудно найти такую переходную форму, которую можно поставить именно в точку расхождения ветвей. Филогенетические схемы было бы правильнее рисовать не в виде деревьев, а в виде отдельных штрихов, между которыми есть разрывы. Все известные переходные формы – "боковые ветви", которые демонстрируют собой стадии эволюции, но не являются непосредственными предками следующего известного крупного таксона.
Например, незадолго до появления четвероногих существовала обширная группа рыб подкласса лопастепёрых, Sarcopterygii, которые по многим признакам сходны с самыми древними земноводными. В ней можно указать группы, которые особенно близки к земноводным (кистепёрые, а среди них – рипидистии, среди рипидистий – остеолепиформы и т. д.), но не удаётся указать группу, которая была непосредственным предком четвероногих. В свою очередь, самых древних амфибий – ихтиостег – никто не рассматривает как предков всех четвероногих. Аналогично в палеонтологической летописи располагаются и следующие стадии эволюции позвоночных вплоть до человека (см. Кэррол, 1992).
Таким образом, переходные формы между крупными таксонами есть, но и разрывы, как будто бы, тоже есть. Это противоречие очень трудно преодолеть, поэтому до сих пор и не утихают споры о сальтационизме. Например, Э. Майр (Майр, 1974) писал о предках человека примерно следующее: австралопитек – "дисгармоничный" тип, он сочетает в себе примитивные и прогрессивные черты, и не может быть непосредственным предком человека, но демонстрирует стадию, которую проходили предки человека, которые пока не найдены. Его противники утверждали, что предки никогда и не будут найдены, потому что новые виды появляется скачкообразно. Ту же ситуацию можно видеть и в вопросе о происхождении рукокрылых, только в более явном виде. Разрыв, который всегда выявляется в палеонтологической летописи при детальном исследовании, в данном случае никак не удаётся скрыть какими-нибудь промежуточными "боковыми ветвями". Если этот разрыв когда-нибудь и будет заполнен палеонтологическими находками, то они будут напоминать или приматов, или шерстокрылов, или других "специализированных" животных, которые в чём-то сходны с рукокрылыми, но никак не являются их предками.
При всём при этом переходные формы не всегда характеризуются адаптивными признаками. У некоторых животных и растений описаны органы, которые имеют какой-то смысл только при максимальном развитии, а первые стадии в их развитии, как будто бы, никакого применения не находят. Такие органы были названы аристогенами (Osborn, 1934) или морфологическими загадками (Willis, 1940). Впервые это явление глубоко проанализировано Г. Осборном (H. Osborn, 1857-1935) на примере эволюции нескольких групп млекопитающих. Так, в позднем кайнозое существовало множество видов титанотериев – млекопитающих, похожих на носорогов. Считается, что хорошо развитые рога дают селективное преимущество их обладателям. Но у предков титанотериев, которые жили достаточно продолжительное время, были только зачатки рогов, небольшие бугорки, которые, как будто бы, совершенно были не нужны им самим, а нужны только их далёким потомкам.
Если кожные складки между конечностями и пальцами у млекопитающих - такие же аристогены, неадаптивные изменения, то значит, вполне возможно существование животных со слабо развитыми крыльями, которые не годятся даже для планирующего полёта, с такими как, например, у дракона из общей книги о животных К. Гесcнера (K. Gessner, 1516-1565) (рис. 2). Что вызвало появление аристогенов? Автор концепции аристогенеза, Г. Осборн считал, что это так же загадочно для нас, как и то, что вызвало рудимент рога на черепе титанотерия.

Рис. 2. Дракон. ("Общая книга о животных" К. Гесснера). Konrad Gesner. Historia Animalum. Book I-IV. 1551-1563. Рисунок взят из: Gesner K. Curious Woodcuts of Fanciful and Real Beasts. A selection of 190 sixteenth-century woodcuts from Gesner’s and Topsell’s natural histories.N. Y. Dover publications, 1971. 111 p. (Рис. На стр. 84)
Происхождение рукокрылых с позиций различных теорий эволюции
Как объяснили бы эти факты сторонники различных эволюционных теорий? Проще всего, наверное, современным сальтационистам, последователям Р. Гольдшмидта (R. Goldschmidt, 1878-1958) или "пунктуалистам". Согласно их представлениям, новые группы живых организмов появились скачкообразно и случайно: у "нормального" млекопитающего вдруг родился детеныш с крыльями, "монстр", который случайно оказался перспективным в эволюции и дал начало обширной группе новых организмов.
Для ламаркистов тоже не составило бы большого труда дать характеристику процесса происхождения рукокрылых. Сам Жан Батист Ламарк (J. B. Lamarck, 1744-1829) анализировал этот случай и не видел в нём ничего особенно загадочного. Наоборот, он приводил его в качестве "весьма убедительного доказательства" действия привычек на форму и состояние органов животных (Ламарк, 1959, С. 432). Так, летающие белки приобрели привычку вытягивать ноги при прыжках, придавая форму парашюта своему телу. Благодаря частому упражнению кожа растянулась на боках и образовала перепонку. У шерстокрылов эта привычка появилась значительно раньше, поэтому и перепонка у них развита сильнее. У рукокрылых эта привычка является ещё более древней, чем у шерстокрылов, поэтому они и достигли полного совершенства в этом отношении. Современные ламаркисты могли бы внести некоторые уточнения по поводу обстоятельств, которые вызвали столь странные привычки – обстоятельства, сложившиеся при жизни в кронах деревьев в лесах с разреженным верхним ярусом. Кроме того, надо особо подчеркнуть длительность этого процесса. Ламарк не был настолько наивным, каким его обычно считают: если в данном случае животное упражняет пальцы, то это не означает, что у него сразу же родится ребёнок с перепонками. Это процесс происходил очень медленно едва заметными шагами.
Согласно дарвинизму, в ходе эволюции предков рукокрылых происходил отбор мелких уклонений по отдельным признакам. Сначала, например, появились небольшие перепонки, затем немного удлинились пальцы или наоборот. Совокупность признаков, необходимых для полёта, появилась очень медленным извилистым путём. Каждый шаг этого пути имел адаптивное значение, что подтверждается примерами из современной фауны. В современной фауне есть животные, демонстрирующие все стадии развития признаков возможных предков рукокрылых (перепонки, удлинение пальцев, удлинение передних конечностей).
Многие дарвинисты добавили бы, что при этом у предков рукокрылых имелась "тенденция" варьировать в определённом направлении или же указали бы на "эффект Болдуина": замену модификаций соответствующими мутациями. Некоторые дарвинисты, очевидно, рассуждали бы о "функциональных задачах", которые организмам поставила среда: переход к древесному образу жизни потребовал развития длинных пальцев, прыжки потребовали развития перепонок и т. д.
С позиции концепций направленной эволюции этот вопрос можно рассмотреть следующим образом. Теоретически можно представить себе несколько направлений в эволюции конечностей: удлинение передних конечностей, удлинение задних конечностей, удлинение пальцев и т. п. Каждое из них реализуется в эволюции млекопитающих. При любом из выбранных направлений возможно появление перепонок, что подтверждается примерами из современной фауны. Таким образом могло сформироваться "определённое направление" в развитии комбинации признаков: удлинение пальцев, удлинение передних конечностей, развитие перепонок. Согласно закону инерции, раз уж эволюционное направление возникло, то оно продолжается до максимально возможного развития, оно развивается подобно росту кристаллов даже если ведёт и в "неадаптивную сторону". Почти повсеместно это направление было обречено на быстрое вымирание, но в некоторых условиях – в лесах типа южно-азиатских или австралийских, или, может быть, в пещерах - сохранилось и развивалось. Эволюционирующие признаки могли быть неадаптивными на каких-то стадиях, но, по крайней мере, не слишком вредным для организма.
Какое из объяснений лучше?
Сальтационизм-пунктуализм в том виде, в котором он сейчас существует, означает отказ от какого бы то ни было объяснения. Появление монстра рассматривается как событие, подобное чуду. В качестве подтверждения его возможности обычно ссылаются на случаи появления жизнеспособных форм с наследственными аномальными признаками. Например, львиный зев с центрально-симметричным цветком, кошки без хвоста, коротконогие овцы и т. п. Но такие аномалии не являются фактами появления новых семейств или классов растений и животных, и все они ничего общего не имеют с "перспективными монстрами" Р. Гольдшмидта. Аномальное строение затрагивает только отдельные признаки, и если считать их аналогией процесса появления рукокрылых, то они доказывают, что могли появиться новые животные или со складками между пальцев, или с удлинёнными передними конечностями, или аномальным строением задних конечностей и т. п., но никак не со всеми этими особенностями одновременно.
Любопытно, что история сальтационизма, по крайней мере, новейшая, никак не связана с исследованиями аномалий, в то время как практически во всех исследованиях аномалий, начиная с Э. Жоффруа Сент-Илера (E. Geoffroy Saint-Hilaire, 1772-1844) или даже Аристотеля, подчёркивались закономерности их появления и утверждалось, что аномалии не могут быть какими угодно<![if !supportFootnotes]>[3]<![endif]>. Таким образом, сальтационная модель должна подкрепляться анализом закономерностей изменчивости или направленности эволюции, что более характерно для иных эволюционных концепций, в корне отличающихся от современного пунктуализма.
Ламаркизм в данном случае кажется устаревшим. Хотя в современной эволюционной биологии о нём нередко вспоминают, но обычно это делается в контексте иммунологии, микробиологии, вирусологии (см. Голубовский, 2000). "Адаптивные мутации", "горизонтальный перенос генетического материала" и другие "неканонические" формы наследственной изменчивости трудно отнести к тем процессам, которые обсуждались выше. Кроме того, ламаркизму (как отчасти и дарвинизму) не соответствует большая скорость эволюции предков рукокрылых. В масштабах геологической истории они появились практически мгновенно.
Дарвинизм и ортогенез дают более убедительные объяснения в общих чертах, хотя в них остаётся нечто не вполне чёткое – "тенденции" или "определённые направления" развития. Некоторые дарвинисты объясняли появление "тенденций" тем, что когда перед организмами средой ставятся новые задачи, то число способов их решения ограничено. Оно может даже быть равным единице, и поэтому создаётся впечатления существования внутренней тенденции развития (см. Гиляров, 1970). Но такая интерпретация закономерностей эволюции всякий раз обвинялась в телеологии. В то же время утверждения "нетелеологических" закономерностей сторонниками идеи направленной эволюции клеймилось сторонниками дарвинизма как мистические, идеалистические, телеологические, предвзятые и т. п. Как ни пытались сторонники ортогенеза утверждать, что они считают "законы эволюции" аналогичными физическим, астрономическим и физическим, за ними прочно утвердилась репутация мистики и ненаучности.
Таким образом, наиболее продуктивные объяснения с позиций различных эволюционных концепций упираются в механизмы "закономерностей" или "тенденций" развития в определённом направлении. Проблема их изучения в наиболее явном виде ставится концепциями направленной эволюции, что отчасти свидетельствует в их пользу. В своё время немало биологов видело главную заслугу дарвинизма именно в том, что он прямо или опосредованно предлагал программы исследований эволюции. Если руководствоваться ортогенезом, то в центре внимания оказываются проблемы закономерностей изменчивости, эволюционных запретов, вопросы нормы и аномалий, морфогенетических корреляций, то есть те вопросы, которые обычно находятся где-то на периферии общей биологии (см. Коваленко & Попов, 1997). Мы выяснили, что некоторые современные программы в данном случае зашли в тупик, то есть не соответствуют поставленным в них задачам. Несмотря на огромное количество основательных и очень дорогостоящих работ по филогении, они ничего убедительного не говорят ни об истории развития группы, ни о её предках. Заметим, что ещё сто лет назад во время бурного развития эволюционной палеонтологии некоторые классики вскоре пришли к выводу о большом вреде склонности "с лихорадочной поспешностью строить бесчисленные генеалогические деревья, гнилые стволы которых, по образному выражению Рютимейера, едва построенные, уже разрушаются, устилая собой почву леса и затрудняя будущие успехи" (Депере, 1921. С. 91). Не ту ли ситуацию мы видим в настоящее время в бурном развитии кладистики?
Выводы
Условия, в которых происходила эволюция ближайших нелетающих предков рукокрылых, можно охарактеризовать вполне определённо. В настоящее время среду, в которой всё живое "стремится в воздух", демонстрируют экваториальные леса типа Южно-Азиатских, то есть те, в которых разрежен верхний древесный ярус. В них многократно происходило появление всевозможных планирующих животных, и вполне вероятно, что в аналогичных условиях появился настоящий полёт. Кроме того, эволюция ближайших предков летучих мышей (одной из двух групп рукокрылых, которые, вероятно, имели разных нелетающих предков) была тесно связана с пещерами. Об этом можно судить по тому, что все они, включая самых древних вымерших, имели высокоразвитую систему эхолокации.
Все признаки из комплекса черт возможных переходных форм от "нормальных" четвероногих млекопитающих к рукокрылым можно видеть по отдельности у различных современных животных. У млекопитающих, которые ведут древесный образ жизни, встречаются и кожные перепонки любой степени развития, и удлинённые пальцы, и гипертрофия передних конечностей. В большинстве случаев перепонки находят какое-то применение при прыжках или планирующем полёте. Это косвенно доказывает жизнеспособность переходных форм между рукокрылыми и "неспециализированными" млекопитающими.
Детали развития совокупности главных отличительных особенностей рукокрылых остаются неизвестными. При попытках его характеризации с позиции различных эволюционных концепций "загадка" сводится к выяснению "тенденций" или "закономерности" изменчивости. Проблема их изучения ставится в наиболее явном виде в концепциях направленной эволюции. Кроме анализа закономерностей изменчивости, в данном случае результативным является использование гипотезы Дж. Виллиса о возрасте-ареале и биогеографическом анализе. По крайней мере, оказалось, что это позволяет ликвидировать некоторые неясности в появлении планирующих животных. В тоже время, сравнительно-филогенетические методы явно зашли в тупик при изучении эволюции рукокрылых.
список литературы
Гиляров М. С. Закономерности приспособлений членистоногих к жизни на суше. М.: Наука, 1970. 276 с.
Голубовский М. Д. Век генетики: эволюция идей и понятий. СПб: СПбФИИЕТ РАН, 2000. 262 с.
Депере Ш. Превращениия животного мира. Петроград: Изд. М. и С. Сабашниковых, 1921. 270 с.
Коваленко Е. Е. & Попов И. Ю. Новый подход к анализу свойств изменчивости // Журнал общей биологии. 1997. Т. 58. No 1. С. 70-83.
Ковтун М. Ф. Морфологический аспект эволюции органов локомоции рукокрылых (Chiroptera) // Современные проблемы эволюционной морфологии. / Отв. ред. Воробьёва Э. И. М.: Наука, 1988. С. 104-117.
Ковтун М. Ф. Проблемы эволюции рукокрылых. // Вестник зоологии. 1990. No 3. С. 3-13.
Константинов А. И. Причины возникновения эхолокации у рукокрылых // Вопросы териологии. Рукокрылые (Chiroptera). М.: Наука, 1980. С. 271-276.
Кэрролл Р. Палеонтология и эволюция позвоночных. М: Мир, 1992. Т. 1-3. 279, 280, 308 с.
Ламарк Ж. Б. Избранные произведения в двух томах. М.: АН СССР, 1955, 1959. 968 с. 895 с.
Майр Э. Популяции, виды и эволюция. М.: Мир, 1974. 460 с.
Панютин К. К. Происхождение полёта рукокрылых. // Вопросы териологии. Рукокрылые (Chiroptera). М.: Наука, 1980. С. 276 – 286.
Попов И. Ю. Монстры в эволюции. // Эволюционная биология: история и теория. СПб: СПбФИИЕТ РАН, 1999. C. 70-81.
Попов И. Ю. & Сендек Д. С. Квинтэссенция эволюции // Эволюционная биология: история и теория. Выпуск 2. СПб: СПбФИЕЕТ РАН, 2003. С. 172-189.
Фауна мира. Млекопитающие. Ред. Соколов B. E. М: Агропромиздат, 1990. 254 с.
Чайковский Ю. В. Элементы эволюционной диатропики. М.: Наука, 1990. С. 62.
Ammerman L. K. & Hillis D. M. A molecular test of bat relationships: monophyly or diphyly? // Systmatic Biology. 1992. V. 41. P. 222-232.
Bocking S. Alpheus Spring Packard and Cave Fauna in the Evolution Debate. // Journal of the History of Biology. 1988. V. 21. No 3. P. 425-456.
Crompton A.W. The enigma of the evolution of mammals // Optima. 1968. 18. N 2. P. 137-151.
Hill J. E. & Smith J. D. Bats. A natural history. Austin: Univ. of Texas Press, 1984. 243 p.
Kovtun M. F. On the origin and evolution of bats. // European bat research. / Eds. Hanбk V., Horбcek I. & Gaisler J. 1987. Praha: Charles Univ. Press, 1989. P. 5-13.
Moore J. & Willmer P. Convergent evolution in invertebrates. // Biological reviews of the Cambridge Philosophical Society. 1997. V. 72. No 1. P. 1-61.
Norberg U. M. Vertebrate flight. Berlin: Springer Verlag, 1990. 291 p.
Novacek M. J. Evidence for echolocation in the oldest known bats. // Nature. 1985. V. 315. P. 140-141.
Osborn H. F. Aristogenesis, the creative principle in the origin of species. // Amer. Nat. 1934. V. 68. P. 193-235.
Padian K. The origins and aerodynamics of flight in extinct vertebrates // Palaeontology. 1985. V. 28. Pt. 3. P. 413-433.
Pettigrew J. D. Flying primates? Megabats have the advanced pathway from eye to midbrain. // Science. 1986. V. 231. P. 1304-1306.
Pettigrew J. Summary of studies on primate phylogeny and "flying primate" hypothesis. // Eds. Racey P. A. & Swift S. M. Symp. Zool. Soc. London. 1999. V. 67. P. 3-26.
Popov I. Yu. El problema de los monstruos en la evoluciуn // Ludus vitalis. 2000. V. VII. Num. 13. P. 35-50.
Rayner J. M. V. The cost of being a bat. // Nature. 1991. V. 350. P. 383-384.
Richards P.W. The tropical rain forest. An ecological study. Cambridge, 1958.
Simmons N. B. The case for chiropteran monophyly. // American Museum Novitates. 1994. No 3103. P. 1-54.
Speakman J. R. & Racey P. A. No cost of echolocation for bats in flight. // Nature. 1991. V. 350. P. 421-423.
Teeling E. C., Scally M., Kao D. J., Romagnoli M. L., Springer M. S., Stanhope M. J. Molecular evidence regarding the origin of echolocation and flight in bats. // Nature. 2000. V. 403. No 6766. P. 188-92.
Vandel A. Biospйologie. La biologie des Animaux Cavernicoles. Paris: Gauthier-Villars Йditeur, 1964. XVIII. 619 p.
Wellnhofer P. Die befiederten Dinosaurier Chinas // Naturwissenschaftliche Rundschau. 2002. No 651. S. 465-477.
Wible J. R. & Novacek M. J. Cranial evidence for the monophyletic origin of bats // American Museum Novitates. 1988. No 2911. P. 1-19.
Willis J. C. Age and Area. Cambridge: Cambridge Univ. Press, 1922. 259 p.
Willis J. C. The Course of Evolution by Differentiation or Divergent Mutation Rather than by Selection. Cambridge: Cambridge Univ. Press, 1940. 207 p.
<![if !supportFootnotes]>
<![if !supportFootnotes]> [1]<![endif]> Что касается третьей группы летающих позвоночных - птерозавров, то о ней сведения ещё менее определённы, чем о птицах и рукокрылых, и она обычно мало обсуждается в связи с анализом происхождения полёта.
<![if !supportFootnotes]> [2]<![endif]> Проблема возможности филогенетических реконструкций по Виллису подробнее анализируется нами в статье "Квинтэссенция эволюции" (Попов & Сендек, 2003).
<![if !supportFootnotes]> [3]<![endif]> История проблемы аномалий в эволюции рассматривается нами подробнее в двух статьях (Попов, 1999; Popov, 2000).