
Проблемы Эволюции |
Санкт-Петербургский государственный университет педагогического мастерства, 2002.
К.В.Галактионов
Санкт-Петербургский государственный
университет педагогического мастерства
Введение
Глава 1. Краткая история развития мегасистематики
Глава 2. Фундаментальные уровни организации живого - прокариоты и эукариоты
Глава 3. Начальные этапы эволюции жизни на Земле
Глава 4. От прокариот к эукариотам
Глава 5. "Воцарение" эукариот
Глава 6. "Уровневый" подход к построению мегасистемы
Глава 7. Попытки построения "филогенетической" мегасистемы
Список литературы
ВВЕДЕНИЕ
Последние три десятилетия наши взгляды на разнообразие живого претерпевают самые что ни на есть драматические изменения. Привычное подразделение живых организмов на растения и животные либо в лучшем случае на растения, грибы и животные, какое лежит и в основе выделения соответственно двух или трех крупнейших таксономических единиц - царств, приходит в конфликт с новыми данными, которые накапливаются по мере совершенствования методического аппарата современной биологии. Они однозначно свидетельствуют, что окружающий нас мир живых существ намного более многообразен и не вмещается в рамки двух-, и даже трех-царственной схемы. В то же время становится совершенно очевидным, что пока еще мы не достигли того уровня осознания проблемы, который позволил бы дать однозначно приемлемую со всех точек зрения метасистему живого. Наши знания по целому ряду групп эукариот еще фрагментарны и появление новых данных сразу же сказывается самым существенным, подчас, образом на построениях таксономистов.
Итак, "момент истины" в мегасистематике пока еще не настал, но он уже не за горами. Развитие представлений о многообразии живых существ перешло ту грань, когда дискуссии, ведущиеся в сугубо специальной литературе, выплескиваются и на более широкую аудиторию, а ряд новых принципов включается в рамки конкретных учебных курсов, соответствующие учебные пособия и обобщающие сводки. Школьный курс, являющийся более консервативным (что, заметим, в целом ряде случаев вполне оправдано), пока еще остается в стороне от новых веяний. Но все же не совсем. Информация о положении дел в мегасистематике обсуждается на страницах журнала "Биология в школе" (Шипунов, 1997; Медников, 1997; Чайковский, 1998 и др.), включается в некоторые пособия для поступающих в вузы (например, Лемеза и др., 1999). В то же время можно констатировать, что в специальной литературе для учителей пока еще отсутствует развернутое описание новых идей по вопросу о происхождении и многообразии живых организмов. Настоящее пособие призвано до некоторой степени заполнить существующий пробел. В его основу легли лекции, которые читались автором на протяжении последних пяти лет на курсах повышения квалификации учителей-биологов и учителей естествознания в СПбУПМ.
Глава 1
КРАТКАЯ ИСТОРИЯ РАЗВИТИЯ МЕГАСИСТЕМАТИКИ
Под термином мегасистематика понимается классификация живых организмов на самом высоком таксономическом уровне, то есть уровне царств (надцарств, подцарств) и выделяемых в последнее время некоторых более высоких таксономических категорий, таких как доминионы и империи. Впервые столь глобальное подразделение живых существ было предпринято Аристотелем (ок. 384-322 до н.э.), который предложил различать растения и животные. Критериями для отнесения живых организмов к той или другой группе служили, согласно Аристотелю, подвижность и чувствительность. Растения неподвижны и нечувствительны, животные - наоборот. Надо отметить, что уже сам Аристотель столкнулся с трудностями при классификации некоторых прикрепленных или малоподвижных организмов, таких как губки, голотурии и асцидии. Он полагал, что в них смешана животная и растительная природа.
Несмотря на эти затруднения, классификация Аристотеля была принята Карлом Линнеем (1707-1778), который в своей знаменитой "Systema Naturae" (1735), выделил два царства: Vegetabilia (растения) и Animalia (животные). Впоследствии к двум критериям различения растительных и животных организмов Аристотеля Жан Батист Ламарк (1744-1829) добавил еще и способ питания - автотрофный для растений и гетеротрофный для животных. Такая двуцарственная система живого просуществовала практически до наших дней, хотя время от времени и подвергалась сомнению.
Осложнения стали накапливаться еще со времени открытия Левенгуком (1632-1723) мира микроскопических организмов, названных им анималькулами. Само название указывало на причисление этих живых существ к царству животных, что основывалось на критерии подвижности. К.Линней в своей системе все известные к тому времени микроскопические организмы объединил в рамках рода Chaos, который включался в отряд Zoophyta, наряду с медузами, головоногими моллюсками, иглокожими и др. Само название Zoophyta указывает на двойственность, по мнению Линнея, природы всех этих живых существ, которых знаменитый шведский натуралист все же относит к царству животных. По мере развития микроскопической техники выяснилось, что среди анималькул есть формы, снабженные хлоропластами, то есть растительные по своей природе, есть питающиеся гетеротрофно, принадлежащие царству животных, и, наконец, имеются отличные от тех и других микроорганизмы (монеры). Всех их Геккель (Haeckel, 1866) предложил выделить в самостоятельное царство Protista (протисты), к которому он также отнес бактерий и некоторых низших многоклеточных животных (губки). Эта точка зрения не нашла поддержки, и в более поздних работах сам Геккель отказался от нее, распределив протистов между царствами растений и животных.
Однако противоречивость двуцарственного подразделения живого становилась все более и более очевидной. Свое выражение это находило в практической систематике, в рамках которой ботаники и зоологи относили (и до сих пор относят!) одни и те же организмы к разным таксонам. К примеру, согласно ботанической номенклатуре эвгленовые выделяются в самостоятельный отдел (тип), динофитовые (динофлагелляты) в подотдел или, по крайней мере, в класс, а вольвоксовые в класс или порядок. Зоологи же рассматривают всех их лишь как отряды в классе жгутиконосцев. Таких "двуцарственных" видов, которые классифицируются одновременно как в качестве растений, так и в качестве животных, насчитывается сейчас около 30 тысяч (Кусакин, Дроздов, 1998). Более того, во многих группах водорослей (криптофитовые, динофитовые, золотистые, зеленые) имеются бесцветные, то есть лишенные хлоропластов формы. Их, следуя критерию способа питания, следовало бы относить к животному царству, хотя их родство с остальными представителями соответствующих таксонов, обладающих хлоропластами, несомненно и никогда ни у кого сомнения не вызывало. Можно вспомнить в этой связи и классический пример некоторых эвгленовых, которые на свету питаются автотрофно, а в темноте переходят к гетеротрофному способу питания.
Несмотря на парадоксальность ситуации, подобная система классификации с молчаливого согласия ботаников и зоологов существовала вплоть до самого недавнего времени, да и поныне еще принята во многих учебных курсах, руководствах и пособиях. Одна из причин подобного положения дел - мелкие размеры простейших живых организмов, что до появления электронного микроскопа резко ограничивало набор признаков, используемых при таксономических построениях.
Ситуация стала постепенно изменяться начиная с 60-х годов, когда в связи с активным внедрением в биологию методов электронной микроскопии (особенно интенсивно эти исследования проводились в 70-80-х годах) стали накапливаться принципиально новые данные по тонкому строению (ультраструктуре) простейших живых организмов. Оказалось, что на этом уровне выявляются достаточно отчетливые морфологические признаки (тонкое строение покровов, жгутикового аппарата, митохондрий, хлоропластов и др.), которые можно использовать в качестве надежных критериев при выяснении степени родства организмов. Другая волна новой информации стала быстро распространяться с 80-х годов со стороны молекулярной биологии, когда появилась возможность сравнивать степень сходства нуклеиновых кислот разных организмов.
При выполнении такого рода исследований при помощи специальных методик проводится так называемый сиквенс, то есть определяется последовательность нуклеотидов в том или ином участке РНК или ДНК. Затем сиквенсы одних и тех же участков нуклеиновых кислот разных организмов сравниваются друг с другом. Число выявленных при этом различий определяет степень родства сравниваемых организмов: чем больше несовпадений в последовательностях нуклеотидов, тем дальше отстоят они друг от друга. Для таксономических построений используются преимущественно рибосомальные РНК (5S рРНК, 16-18S рРНК, 23-28S рРНК), в ряде случаев митохондриальная ДНК и последовательности ядерной ДНК, кодирующие определенные белки (а- и b- глобулины, стрессовые или хит-шоковые белки, цитохромы и др.).
Сразу следует оговориться, что считать молекулярно-биологические данные истиной в последней инстанции и панацеей от всех бед, что нередко практикуется, нельзя ни в коем случае. Все эти построения зиждутся на признании, в той или иной форме, гипотезы о нейтральности эволюции и следующего из нее принципа молекулярных часов (Кимура, 1985). Согласно последнему, замены нуклеотидов (то есть точечные мутации) в ходе эволюции происходят с постоянной частотой, одинаковой для разных групп организмов. Это-то и дает основание для определения степени родства между ними при сравнении сиквенсов гомологичных участков нуклеиновых кислот. Более того, молекулярные часы можно калибровать. Для этого достаточно вычислить число замен нуклеотидов в определенных участках нуклеиновых кислот двух организмов (или большего их числа), время дивергенции которых известно из палеонтологической летописи. Затем результаты такой калибровки можно переносить на любые другие группы организмов.
В действительности скорость эволюции, а соответственно и скорость накопления мутаций, не остается постоянной даже в пределах одной филогенетической линии (Татаринков, 1987; Тыщенко, 1992). Периоды интенсивного протекания эволюционных изменений сменяют, как правило, длительные периоды застоя (стазиса). Эти представления учитываются в современных молекулярных работах. Для анализа выбирают, как правило, более-менее консервативные типы или участки нуклеиновых кислот, параллельно используются результаты сопоставления сиквенсов разных типов или участков ДНК и/или РНК. Несмотря на это, к молекулярно-биологическим данным следует относится с известной осторожностью и при их интерпретации учитывать традиционные морфологические признаки.
Так или иначе, но именно ультраструктурные и молекулярно-биологические данные привели к той революции в мегасистематике живого, которую мы переживаем в настоящее время. Во-первых, было выявлено несомненное родство между группами, родственные связи между которыми ранее даже не предугадывались. Так. например, среди водорослей эвгленовые оказались близкородственны группе паразитических жгутиконосцев-кинетопластид (к ним, в том числе, относятся трипанозомы - возбудители таких опасных заболеваний, как сонная болезнь, болезнь Чагоса и др.), а динофитовые - инфузориям и споровикам, всегда считавшимися зоологическими объектами и не обнаруживавшими на светооптическом уоовне ни малейшего сходства как с динофитовыми, так и между собой Во-вторых, различия между представителями пазных групп протистов оказались, по крайней мере, не меньше, а зачастую и больше, чем между типичными растениями и животными.
Наметился кризис в классических представлениях о мега-системе живого (Кусакин, 1995), который до определенной степени напоминает создавшийся в физике 20-х годов вслед за открытием новых свойств электрона. Путь к его разрешению видится в создании многоцарственных систем живых организмов, различные варианты которых можно найти в сводках последних десятилетий (Whittaker, 1969; Cavalier-Smith, 1981, 1999; Corlis, 1987; Старобогатов, 1986; Серавин, 1989; Карпов, 199о! 2000; Кусакин, 1995; Кусакин, Дроздов, 1998 и др.). Однако, прежде чем приступить к их анализу, следует обсудить некоторые общие принципы, на которых может базироваться современная мегасистема живого.
Глава 2 ФУНДАМЕНТАЛЬНЫЕ УРОВНИ
ОРГАНИЗАЦИИ ЖИВОГО -ПРОКАРИОТЫ И ЭУКАРИОТЫ
Прежде всего требует низвержения старая парадигма, согласно которой мир живых существ достаточно отчетливо подразделяется на растения и животные. Подобная классификация слишком тесна для того реального многообразия живого, которое вырисовывается современной наукой, и, образно говоря, представляет собой не более чем черно-белое изображение действительности. А коль скоро подобное подразделение не может считаться адекватно соответствующим новым данным, то следует искать иные, чем использовавшиеся ранее, критерии для выделения крупнейших таксономических рангов живых существ. При этом могут быть задействованы два различных методологических подхода, условно обозначаемых как "уровневый" и "филогенетический". Первый предполагает выделение неких масштабных этапов эволюции, каждый из которых отражает определенный уровень структурной организации живого, контрастно отличающийся от остальных. Второй направлен на поиск родственных связей между живыми организмами и выделение их группировок, объединенных происхождением от обшей предковой формы. Таким образом, если уровневый подход можно представить как горизонтальный срез эволюции, то филогенетический отражает попытку выстраивания некоторых вертикальных линий, причем уровень организации организмов, стоящих у их основания и у вершины, может существенно отличаться.
Попытаемся вначале использовать возможности уровнево-го подхода. Сразу приходят на ум те два фундаментачьных уровня организации живых систем, на существование которых впервые обратил внимание Геккель (Haeckel, 1878). Он выделил группу безъядерных протистов - монер (бактерии), тело которых, в соответствии с его представлениями, еще не поделено на протоплазму и ядро. Впоследствии французский ученый Шаттон (Chatton, 1925 - цит. по Серавин, 1991) предложил различать среди всех живых существ две группы. Это Prokaryota (от греч. pro -- до, прежде и karyon - ядро), прокариоты или доядерные, то есть не имеющие истинного ядра, и Eukaryota (от греч. eu - истинный, полный и karyon - ядро), эукариоты (эвкариоты), обладающие истинным ядром. Предложенное французским исследователем подразделение, сделанное еще задолго до эры электронной микроскопии и открытия роли нуклеиновых кислот как носителей генетической информации, в дальнейшем блестяще подтвердилось и в настоящее время общепризнанно.
К прокариотам относятся все бактерии, включая цианобактерий, которых до сих пор еще зачастую именуют сине-зелеными водорослями. Это наиболее просто устроенные организмы, размеры которых ограничены несколькими микронами. Снаружи бактерии, как правило, одеты плотной клеточной стенкой, под которой располагается наружная мембрана. Роль ядра играет нуклеоид, в состав которого входит кольцевая нить ДНК, закрепленная на внутренней стороне наружной мембраны. Нуклеоид лишен оболочки и не обладает ядрышком (Маргелис, 1983; Громов, 1985; Серавин, 1986, 1991 и др.). Перераспределение генетического материала в ходе бесполого и полового (рекомбинация) размножения бактерий не связано с механизмами митоза и мейоза. Для генов бактерий не характерна интрон-экзонная структура (исключение - архебактерии, речь о которых ниже), которая свойственна генам эукариот. Соответственно, отсутствует у них и сплайсинг (см. ниже).
Структурные гены, кодирующие ферменты, необходимые для протекания того или иного биохимического процесса, в геноме бактерий тесно сцеплены друг с другом и с участком ДНК, контролирующим их транскрипцию - оператором (рис. 1). Все вместе они образуют единицу генетического материала, получившую название оперон (см. Албертс и др., 1986; Инге-Вечтомов, 1989). Оператор состоит из небольшого числа нуклеотидов (21 для лактозного оперона) и перекрывается с расположенным перед сцепленными структурными генами участком ДНК, называемым промотором. Последний служит участком связывания РНК-полимеразы, фермента, необходимого для транскрипции ПНК Непременным компонентом оперонной регуляции явтяется ген регулятор. Он кодирует белок-репрессор, который имеет сродство к оператору. Свободный репрессор соединяется с оператором и тем самым препятствует посадки на промотор РНК-полимеразы. Считывание информации со структурных генов оперона оказывается в этом случае невозможным. Однако, если в бактериоплазме присутствует специфичное для каждого репрессора вещество - индуктор, то оно взаимодействует с репрессором, в результате чего последний теряет способность связываться с оператором. Соответственно, оперон "открывается" и происходит транскрипция его структурных генов. Как правило, роль индуктора играет вещество, которое служит исходным субстратом для ферментов, кодируемых тем или иным опероном. Механизмы регуляции транскрипции оперонов весьма различны, но оперонный принцип организации генетического материала характерен для бактерий и практически не встречается у эукариот.

Содержимое тела бактерий не компартментализовано, то есть в нем нет отграниченных собственными цитомембранами органелл - непременной черты организации эукариотной клетки (Громов, 1985; Серавин, 1986, 1991; Заварзин и др., 1992). В бактериоплазме отсутствуют митохондрии, хлоропласты, центриоли (кинетосомы), диктиосмы комплекса Гольджи, лизосомы и др. В связи с этим ассамблеи ферментов, сосредоточенные в клетках эукариот внутри органелл, у бактерий приурочены к поверхностной мембране и ее временным или постоянным инвагинациям (впячиваниям внутрь бактериоплазмы) - мезосомам. Последние, являясь непосредственным продолжением поверхностной мембраны, считаться цитомембранами в том смысле, в котором этот термин используется для обозначения мембран, окружающих органеллы эукариот-ных клеток (см. ниже), не могут. Рибосомы бактерий отличаются от эукариотных по числу белков и коэффициенту седиментации - 70S, в отличие от характерной для рибосом эукариот 80S. Имеются различия и в химическом составе мембран по белкам, фосфолипидам, жирным кислотам и др. Важной характерологической чертой организации прокариот является и отсутствие в бактериоплазме структурного матрикса - цитоскелета, который у эукариот представлен системами микрофиламентов (их образуют разные белки, в том числе актин, миозин, нексин и мн. др.), микротрубочек (состоящих из белка тубулина) и гетерогенной по белковому составу системой промежуточных филаментов (Заварзин и др., 1992). Остается добавить, что жгутик, имеющийся у ряда бактерий, не имеет ничего общего ни по строению, ни по особенностям функционирования с одноименной органеллой эукариот (Громов, 1985).
Среди прокариотных организмов выделяются два основных ствола - эубактерии и архебактерии. Своеобразие последних было выявлено относительно недавно (см. обзор Куса-кин, Дроздов, 1998). Архебактерии образуют небольшую группу (немногим более 45 видов) прокариот, живущих в экстремальных условиях: метанообразующие, галобактерии и др. От остальных бактерий (эубактерии) их отличают особенности структуры рибосомальных 5S рРНК, 16S рРНК и транспортных РНК, а также химический состав клеточной стенки и наружной мембраны. Последняя однослойна, то есть состоит из одного слоя липидов, а не из двух, как у эубактерии и эукариот. Кроме того, для генов архебактерии характерна интрон-эк-зонная, или мозаичная структура (см. ниже), которая раньше считалась неотъемлемой чертой организации эукариот и не встречается у эубактерии. Ее обнаружение у генов архебактерии позволяет предположить, что мозаичная структура генов была присуща самым древним живым организмам, а не возникла de novo при переходе от прокариот к эукариотам (Георгиев, 1989).
Принципиально иной характер морфо-функциональной организации присущ клетке эукариот (см. Албертс и др., 1986; Серавин, 1986, 1991; Заварзин и др., 1992). Сразу оговоримся, что отличия ее от прокариот далеко не исчерпываются только наличием структурно оформленного ядра, которое, конечно, является непременным атрибутом каждой эукариотной клетки. Ядро одето ядерной оболочкой, состоящей из наружной и внутренней мембран, которую пронизывают сложно устроенные ядерные поры. В них располагаются белковые структуры порового комплекса, состоящие из упорядоченно ориентированных глобул. Собственно генетический аппарат представлен нитями ДНК, которые при помощи комплекса белков-гистонов укладываются в хромосомы. Характер упаковки ДНК меняется в ходе клеточного цикла. Как четко оформленные образования, видимые даже при наблюдении в световой микроскоп, хромосомы становятся заметными только на определенных этапах митоза или мейоза. В это время происходит компактизация укладки комплекса ДНК-гистоны, благодаря формированию дополнительных петель и спиралей нескольких уровней. В остальное время (интерфаза) хромосомы находятся в деспирализованном состоянии и их совокупность в ядре обозначается как хроматин. В ядре имеется также и матрикс, который составлен преимущественно белками (в том числе и актином) и выполняет в основном скелетообразующую функцию.
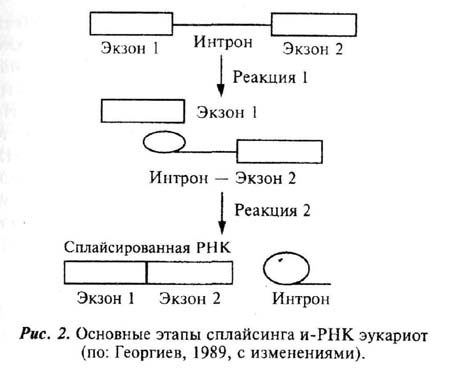
Для генов эукариот, как уже отмечалась выше, характерна мозаичная структура (см. Георгиев, 1989; Инге-Вечтомов, 1989). Это подразумевает, что они складываются из отдельных блоков, одни из которых - экзоны - несут информацию о составе кодируемой данным геном белковой молекулы, другие же - интроны - таковой не несут и отделяют экзоны друг от друга. В ходе транскрипции ядерной ДНК образуется иРНК, на которой записаны как экзонные, так и интронные участки того или иного гена (рис.2). Эта незрелая иРНК входит в процесс, получивший название сплайсинга, в ходе которого ненужные участки молекулы (интроны) вырезаются, а оставшиеся (экзоны) сшиваются конец в конец. Зрелая иРНК, состоящая только из сшитых друг с другом экзонных участков, поступает через ядерные поры в цитоплазму.
Регуляция транскрипции по типу опероыа для эукариот не характерна (Георгиев, 1989; Инге-Вечтомов, 1989). Каждый структурный ген в составе хромосомы имеет свой промотор (участок связывания РНК-полимеразы) и способен транскрибироваться самостоятельно. Для активации этого процесса обычно требуется регуляторный белок, который присоединяется к промотору и тем самым облегчает "посадку" на промотор РНК-полимеразы. Подобный тип регуляции, когда регуляторный белок служит для открытия, а не для запрещения транскрипции, обозначается как позитивная регуляция. Она широко распространена у эукариот, тогда как у прокариот чаще встречается негативный вариант (регуляторный белок блокирует транскрипцию), как в разобранном выше случае экспрессии бактериального оперо-на. Несмотря на отсутствие оперонного принципа в организации генетического материала эукариот, у них имеет место координация транскрипции даже несцепленных генов. Один из возможных механизмов координированной экспрессии определенных групп генов заключается в наличии генов-регуляторов, кодирующих так называемые ключевые белки-регуляторы (Албертс и др., 1986). Эти белки способны активизировать не один, а целую группу генов, тем самым обеспечивая в конечном итоге одновременное производство различных белковых молекул. Важными регуляторными элементами транскрипции у эукариот служат и последовательности, названные энхансерами и сайленсерами, или глушителями. Первые повышают эффективность транскрипции, вторые, наоборот, ее прерывают. Эти последовательности могут находиться за сотни и даже тысячи пар нуклеотидов от регулируемого гена. Механизм их воздействия на транскрипционную активность связан с изменением структуры хроматина (Георгиев, 1989; Инге-Вечтомов, 1989). Наконец, экспрессии генов эукариот могут регулироваться и после транскрипции (например, в ходе сплайсинга и-РНК), и в процессе трансляции (например, исключение возможности использования определенной и-РНК в качестве матрицы для синтеза белка, хотя она и присутствует в цитоплазме) и после нее (например, на уровне ферментов, необходимых для активации вновь синтезированного белка).
Поверхностная (плазматическая) и внутренние (цито-мембраны) мембраны эукариотной клетки существенно отличаются от бактериомембраны по своему биохимическому составу. Важнейшей их особенностью является способность к везикуляции, то есть к замыканию с образованием разного размера пузырьков, вакуолей и цистерн (Серавин, 1986, 1991). Кроме того, мембраны одного типа способны сливаться друг с другом (см. ниже). В цитоплазме имеется развитый цитоскелет, который, как уже упоминалось выше, построен из актиновых, миозиновых и иных микрофиламентов и из микротрубочек. Помимо поддержания формы клеток, цитоскелет, благодаря механохимическим свойствам входящих в его состав белковых систем (актин-миозин, тубулин-динеин и др.), обеспечивает перемещение в цитоплазме органелл и везикул разной природы, все процессы, связанные с эндоцитозом (фагоцитоз, пиноцитоз) и экзоцитозом (транспорт заключенных в мембранные пузырьки веществ из клетки наружу), амебоидное и иные способы клеточного движения.
Наконец, в клетке эукариот всегда имеется несколько компартментов. Это органеллы, входящие в состав практически всех клеток эукариотных организмов, независимо от их положения на эволюционной лестнице. К их числу, помимо ядра, относятся митохондрии, хлоропласты, комплекс Гольджи, эндоплазматический ретикулюм (ЭПР), лизосомы и др. Кроме того, у ряда специализированных клеток, прежде всего протистов, имеются и особые органеллы, такие как сократительные и пищеварительные вакуоли, экструсомы, кинетопласты и многие другие. Ничего подобного в клетках бактерий никогда не встречается. Важно отметить, что имеются определенные различия в химическом составе плазматической мембраны и цитомембран, отграничивающих различные клеточные компартменты. Это делает невозможным их прямое слияние друг другом. Предельно упрощая ситуацию, можно сказать, что в ходе экзоцитоза секреторная гранула не сможет встроиться в наружную плазматическую мембрану, если она будет окружена цитомембраной, характерной, например, для ЭПР. "Переупаковка" синтезируемых клеткой продуктов - это одна из функций комплекса Гольджи. В его уплощенные цистерны (диктиосмы) поступают по цистернам ЭПР синтезированные клеткой белки. Здесь они упаковываются в пузырьки, одетые мембраной, которая по своему составу аналогична плазматической. Такие пузырьки отрываются от диктиосом и транспортируются при помощи цитоскелетных структур в периферические участки цитоплазмы. Здесь окружающая их мембрана встраивается в наружную плазматическую, а содержимое пузырька при этом изливается наружу.
Столь значимая разница в организации прокариот и эукариотной клетки привела Л.Н.Серавина (1991) к выводу, что бактерия по уровню своей структурной и функциональной организации еще не является клеткой. Это безъядерный, доклеточный организм, который Л.Н. Серавин (1991) предлагает именовать пробитом (от лат. pro - до, прежде и cytus - клетка). В нем выделяются три основных компонента: бактериомембрана, нуклеоид и бактериоплазма, причем каждый из этих компонентов не является прямым гомологом таких частей эукариотной клетки (эуцита по Л.Н. Серавину), как, соответственно, наружная плазматическая мембрана, ядро и цитоплазма (Серавин, 1991). Чтобы разобраться, каким путем произошло формирование двух столь принципиально различных уровней организации живых организмов, попытаемся вкратце охарактеризовать основные этапы ранней эволюции жизни на Земле.
Глава 3
НАЧАЛЬНЫЕ ЭТАПЫ ЭВОЛЮЦИИ ЖИЗНИ НА ЗЕМЛЕ
Сразу стоит оговориться, что при любой попытке реконструкции начальных этапов становления живых организмов мы становимся на достаточно зыбкую почву. Все дело в том, что тектонические процессы, извержения вулканов, эрозия полностью уничтожили все следы тех событий, которые отделяет от нас несколько миллиардов лет. Поэтому даже если когда-нибудь каким-то образом удастся получить "жизнь в пробирке", то никто не сможет утверждать, что развитие жизни на нашей планете шло именно по такому сценарию. Нам остается возможность определить один (или несколько) вероятных путей, исходя из современного уровня знаний.
Другой момент, который следует предварительно отметить, это то, что все события, связанные с возникновением жизни, протекали сотни миллионов лет, срок, который трудно осознать, учитывая, что, к примеру, вся история Человека разумного (Homo sapiens) насчитывает не более полумиллиона лет. Поэтому вряд ли можно согласиться с получившим широкую известность высказыванием астронома Ф.Хойла о том, что мысль о возникновении жизни в результате случайных взаимодействий молекул столь же нелепа и неправдоподобна, как и утверждение, что ураган, пронесшийся над мусорной свалкой, может привести к созданию "Боинга-747" (пит. по: Грин и др., 1990). Действительно, если рассматривать возникновение и развитие жизни как единомоментный процесс, то с мнением Хойла невозможно не согласиться. Однако даже самые невероятные с позиций наших сегодняшних знаний события, по-видимому, могли произойти на временном отрезке длительностью в миллионы лет. Кроме того, как справедливо указывает Хорган (1991), следует учитывать, что результат, сам по себе чрезвычайно маловероятный, может быть достигнут посредством череды последовательных промежуточных событий, каждое из которых само по себе вполне осуществимо. Исходя из этих посылок, мы и попытаемся обозначить основные этапы ранней эволюции жизни.
Первые шаги биопоэза (процесс перехода от неживого к живому) принято обозначать как период химической эволюции (Дикерсон, 1981). Начался он вскоре после возникновения Земли из пылевого облака, каковое событие произошло 4 6 млрд. лет назад. По-видимому, примерно 4,5-4,4 млрд. лет назад могли протекать начальные этапы химической эволюции жизни (Фолсом, 1982; Chang, 1999). Это было время активного вулканического газовыделения. Вода в виде паров выделялась из гидратированных минералов. Она конденсировалась и выпадала в виде сильных, почти непрерывных ливней. В результате образовывались неглубокие водоемы и пруды, в которые с дождями попадал вулканический пепел. Одной из важнейших предпосылок для возникновения жизни мог послужить восстановительный характер атмосферы, на что еще в 1929 г. указал один из основоположников гипотезы о развитии жизни естественным путем - английский биохимик Дж.Холдейн. В этой первичной атмосфере отсутствовал кислород, накопление которого проходило значительно позднее и почти исключительно биогенным путем, то есть в результате жизнедеятельности фотосинтезирующих организмов. Отсутствие такого мощного окислителя, как кислород, позволяло спонтанно образовывавшимся органическим соединениям аккумулироваться в воде. Помимо того, до появления в атмосфере кислорода в ней не было того тонкого слоя озона, который, располагаясь на высоте около 25 км, поглощает в наши дни большую часть ультрафиолетового излучения Солнца. Ультрафиолетовые лучи свободно достигали поверхности и поставляли энергию, способную обеспечить синтез органических веществ из воды, двуокиси углерода, метана, аммиака и водорода, которые, по-видимому, присутствовали в первичной атмосфере. Другим источником энергии могли быть грозовые разряды.
Такого рода условия были смоделированы в получивших широкую известность опытах С. Миллера, описание которых вошло во многие учебники. В запаянной колбе через газовую смесь аммиака, метана и водорода в присутствии паров воды пропускались электрические разряды. Через несколько дней на стенках колбы и на воде появился красноватый налет, который, как оказалось, содержал ряд аминокислот. С 1953 г., то есть с момента постановки Миллером своего опыта, подобные эксперименты воспроизводились во многих лабораториях мира, причем использовались как различные газовые смеси, так и источники энергии. Результат всегда был один: если исходная смесь содержит углерод (в составе какого-либо газа), азот, воду и немного водорода, то на выходе образуется широкий спектр простых органических соединений и даже полимеры. В числе этих соединений были выявлены не только аминокислоты, но и углеводороды, предшественники Сахаров, основания нуклеиновых кислот, жирные кислоты и реакционноспособные промежуточные соединения (Фолсом, 1983). Возникли представления о так называемом "холдейнском" или первичном бульоне, т.е. насыщенном простыми органическими соединениями первичном океане.
Итак, первый этап химической эволюции - образование простых органических соединений из неорганических в условиях древней Земли - оказался вполне воспроизводимым.
Несколько сложнее обстоят дела со следующим этапом, которым, следуя логике, должен был стать этап появления полимерных соединений. Трудность здесь заключается в том, что необходимо уяснить, как в условиях первичного океана могли протекать реакции, требующие одновременно притока энергии и отщепления воды. При избытке воды реакция синтеза полимера сдвигается влево, то есть в сторону гидролиза образовавшегося полимерного соединения. Гидролиз сопровождается выделением свободной энергии и, соответственно, может происходить спонтанно. Для сдвига же реакции вправо, то есть в сторону полимеризации, требуется либо повысить концентрацию реагентов и добиться удаления воды, либо обеспечить сопряжение данного процесса с какой-либо реакцией, сопровождающейся выделением энергии, что позволило бы довести полимеризацию до конца. Обе эти возможности исследуются химиками, пытающимися решить загадку предбиологического синтеза полимеров (Дикерсон, 1981; Фолсом, 1983; Chang, 1993).
В современных биологических системах реакции полимеризации, как известно, осуществляются при участии АТФ, служащей источником энергии, и ферментов, которые сопрягают реакции, потребляющие энергию, с реакциями, служащими ее источником. Подобная схема - результат достаточно длительной эволюции и не могла реализовываться в предбио-логическое время. Тем не менее, нельзя исключить возможности активации мономеров, благодаря реакции с соединениями, обладающими большим количеством свободной энергии (карбодиимиды, фосфаты и др.). Активированные таким образом мономеры могли взаимодействовать друг с другом с образованием полимерных соединений (рис. 3).

Другой возможный вариант, который сразу приходит на ум при анализе проблемы удаления воды, это вероятность протекания предбиологического синтеза полимеров в мелких лужицах, у уреза воды, где в результате испарения могли бы создаваться высокие концентрации органических веществ. Однако многие важные предшественники биополимеров, такие как цианистый водород, формальдегид, аммиак и др., сами летучи и не могут концентрироваться подобным образом. Весьма интересным в этой связи выглядит предположение Дж. Бернала о концентрировании молекул на поверхности таких минералов, как слюды и глины (Дикерсон, 1981). Так, глины образуют упакованные стопки силикатных пластин, между которыми имеются прослойки воды. Благодаря слоям воды обе стороны прилежащих друг к другу пластин становятся доступными для молекул, диффундирующих в толщу глин, что чрезвычайно увеличивает общую адсорбционную поверхность. Кроме того, присутствие в силикатных пластинах положительно и отрицательно заряженных ионов может способствовать связыванию заряженных молекул и даже формировать примитивные каталитические центры для некоторых реакций. Возможность протекания полимеризации ряда аминокислот в присутствии определенных минералов нашла экспериментальное подтверждение.
Традиционно следующим этапом в развитии жизни на Земле принято считать формирование протобионтов. Действительно, любое живое существо всегда отделено от окружающей среды определенной поверхностью раздела. Поэтому и проблема поиска возможных вариантов появления такого рода отдельностей долгое время приковывало к себе внимание исследователей. Пионером в этой области заслуженно считается А.И.Опарин, который вместе со своими сотрудниками на протяжении многих лет экспериментировал с коацерватами, то есть обогащенными полимером коллоидными каплями, взвешенными в среде, богатой водой (Опарин, 1966). Эти капли образуются в смесях полимеров, имеют размер от 1 до 500 мкм и отделены от внешней среды более плотным наружным слоем, который с известной натяжкой можно считать прообразом клеточной мембраны. Капли эти крайне нестабильны, но могут стабилизироваться, если в них экспериментально вызывают своего рода примитивный метаболизм. Например, в одном из опытов А.И. Опарина было показано, что если в среду, содержащую коацерваты, добавить фермент фосфорилазу, то он концентрируется внутри капель. При добавлении к окружающей такие коацерваты среде глюкозо-1-фосфата он диффундирует в капли и, полимеризуясь под действием фермента, превращается здесь в крахмал. Образующийся крахмал увеличивает объем капли, она растет и при достижении определенного размера распадается на несколько дочерних капель.
Другая модель протобионта была предложена С. Фоксом. Им было показано, что если нагретую смесь аминокислот поместить в воду, то образуются белковоподобные молекулы со случайной последовательностью аминокислот - протеиноиды. В воде протеиноиды самособираются в микросферы диаметром 1-2 мкм. Они одеты толстой оболочкой, состоящей из частично гидрофобных протеиноидов (Фолсом, 1982). Выяснилось, что микросферы могут катализировать некоторые химические реакции, например, расщепление глюкозы. Фокс предложил рассматривать микросферы как первичные клетки, которые подвергались отбору в зависимости от их каталитической активности, обеспечивающей первичный метаболизм. Они образовывались в условиях древней Земли, по мнению Фокса, из аминокислот, которые могли концентрироваться в испаряющихся водоемах, полимеризоваться под действием тепла (например, на конусах горячего вулканического пепла) и самособираться в микросферы при смыве дождем.
В моделях Опарина и Фокса было продемонстрировано, что образование коацерватов или микросфер типично для полимеров в растворе. Их шансы на "выживание" повышаются, если они способны катализировать какие-либо простые реакции, в результате которых увеличивается их объем или возрастает прочность барьера, отделяющего от внешней среды (Дикерсон, 1981), Можно представить, что такие капли в громадном числе образовывались в первичных водоемах, наполненных органическими полимерами. В ходе длительного химического отбора могли сохраняться капли, способные извлекать из внешней среды энергию и вещество и включать их в соединения, которые обеспечивают "выживаемость" капель. Когда капли становились большими, они распадались на дочерние капли. Явно имеет место некоторая имитация процессов, проходящих в живых системах. За одним существенным "но": отсутствует механизм, который бы гарантировал, что все дочерние капли получат полный набор катализаторов, которым обладала материнская капля. Иными словами, нет генетического аппарата.
У любых современных организмов имеются две главные особенности: способность к метаболизму, обеспечивающему выживание особи, и к репродукции, обеспечивающей выживание в ряду поколений. Модели Опарина-Фокса и им подобные демонстрируют возможный путь эволюционного становления только первой из этих характерных черт живого. Поэтому не удивительно, что многие исследователи пытались подойти к проблеме становления протоклеток с другой стороны, то есть представить эволюцию генетического аппарата. Широкую известность в этой связи получила концепция английского ученого Р.Докинза (1989), обнародованная им в конце 70-х гг. Вся история, по Докинзу, началась с появления в первичном океане некой замечательной молекулы, обозначенной им как репликатор. Название говорит за себя - эта необычная молекула была способна реплицироваться, то есть производить копии самой себя. Строительные блоки, необходимые репликатору для самовоспроизведения, брались прямо из окружающего первичного океана. При копировании неизбежно возникали ошибки, что привело к появлению различающихся друг от друга репликаторов. А если есть изменчивость, то возможен и отбор, который мог быть направлен на повышение стабильности молекул, увеличение скорости их воспроизведения и т.п. На определенном этапе эволюции запас строительных блоков в океане стал истощаться, и между репликаторами появилась конкуренция. Некоторые из них, возможно, приобрели способность химически разрушать других репликаторов и использовать их строительные блоки для воссоздания самих себя. Другие же нашли способ защитить себя, отгородившись белковой стенкой. Так возникли первые "машины выживания", обеспечивавшие сохранность заключенных в них репликаторов. Дальнейшая эволюция шла по пути совершенствования "машин выживания" и привела к тому биоразнообразию, каковое в настоящее время имеется на планете. Соответственно, все живые существа, населяющие Землю, есть ни что иное, как "машины выживания" "эгоистичных" генов. Несмотря на некоторую экстравагантность, в гипотезе Докинза достаточно четко обозначены основные этапы эволюции: появление самореплицирующихся молекул (читай - ДНК) и последующее возникновение систем биосинтеза белка.
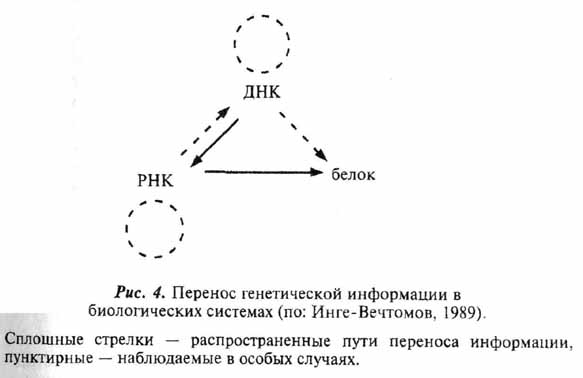
Все концепции такого рода наталкивались на одну, казавшуюся непреодолимой, трудность. Суть ее легко понять, если вспомнить центральную догму молекулярной биологии, сформулированную еще Ф. Криком (рис. 4). Согласно ей, информация воспроизводится при репликации ДНК, переносится от ДНК к РНК и реализуется в виде белка. При этом и репликация, и транскрипция, и трансляция обеспечиваются специфичными ферментами, то есть белками. Таким образом, для нормального функционирования генетического аппарата нужны белки, а для синтеза белков нужна информация, записанная на ДНК, и сложный механизм ее реализации. Встает знакомый вопрос: что было раньше, курица или яйцо?
Ответ на него стал просматриваться в 80-е годы, когда американскими учеными Т. Чеком и С. Альтманом были открыты РНК, которые могли действовать как катализаторы в реакции сплайсинга (Хорган, 1991). Таким образом, было показано, что РНК способны к выполнению ряда функций белков, включая каталитическую активность. А коль скоро РНК может служить ферментом, то вполне вероятно ее самовоспроизведение, то есть она может быть и "курицей" и "яйцом" одновременно. Это сразу наводит на мысль, что именно РНК, а не ДНК должны претендовать на роль Докинзовского первичного репликатора. В пользу того, что именно РНК могли представлять первичный генетический материал, говорит и ряд других фактов, обобщенных недавно А.С.Спириным (2001): в химическом синтезе и в биохимических реакциях ри-бонуклеотиды предшествуют дезоксирибонуклеотидам, последние являются продуктами модификации первых (1); в самых древних, универсальных процессах метаболизма широко пред-славлены рибонуклеотиды, но не дезоксирибонуклеотиды, включая основные энергетические носители типа АТФ и др. (2), репликация РНК может происходить без какого-либо участия ДНК, а для редупликации ДНК необходимо участие РНК-затравки в инициации синтеза цепи ДНК (3).
Все это привело к формулированию гипотезы о "мире РНК", впервые высказанной в 1986 г. У. Гильбертом (Gilbert, 1986). Она предполагает, что важнейшим этапом в становлении жизни стало появление простых самовоспроизводящихся молекул РНК. Это был первый самодостаточный биологический мир, поскольку молекулы РНК функционировали в нем и как генетический материал, и как катализаторы при собственном воспроизведении (Orgel, 1998). В ходе эволюции на основе мира РНК должно было происходить становление механизмов биосинтеза белка, обособление белоксинтезирующих систем от среды (возможно, в виде коацерватных капель) и, наконец, передача функции носителя генетической информации от РНК к ДНК. Последняя, обладая двухцепочечной структурой, более надежна в этом отношении, чем РНК.
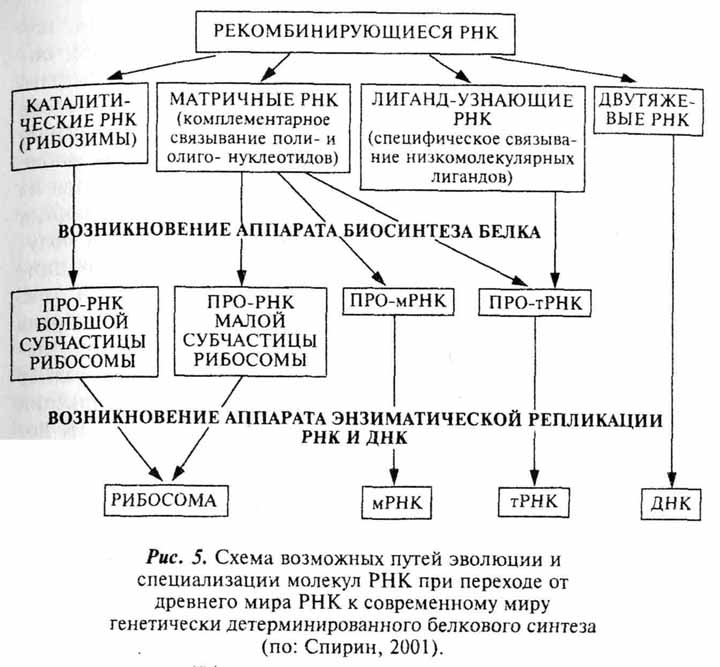
Что касается конкретных путей реализации вышеприведенной схемы, то тут больше вопросов, чем ответов. На сегодняшний день не существует удовлетворительного объяснения тому, как синтез полипентидов и белокподобных соединений мог быть сопряжен с РНК и подпасть под генетический контроль (Спирин, 2001). Согласно предположению А.С. Спирина, предпосылкой для такого события могло стать наличие значительного разнообразия РНК. Первичный аппарат синтеза белка мог включать прo-рибосомальныеРНК, про-иРНК и про-тРНК. Такая система могла бы синтезировать ряд первичных ферментов, в том числе и НК-полимеразы, обеспечивающие полимеризацию нуклеотидов. На основе этих энзимов формировался тот механизм воспроизведения НК, который характерен для современных живых систем (рис. 5).
Концепция мира РНК, которая в настоящее время практически общепризнанна, далеко не так безупречна, как бы того хотелось. Помимо уже обсуждавшейся выше проблемы сопряжения синтеза белка с РНК, остается открытым вопрос о путях предбиологического синтеза РНК (искусственный синтез РНК и составляющих ее нуклеотидов проходит с большим трудом даже в оптимальных условиях эксперимента), подвергается сомнению сама возможность ауторепликации РНК вне контролируемых экспериментатором условий опыта и др. Эти сложности породили значительное число концепций, предполагающих, что миру РНК предшествовал мир иных, более просто устроенных репликаторов, которые как бы "подготовили почву" для органической жизни (Хорган, 1991).
Наиболее широкую известность среди них получила, пожалуй, гипотеза глиняного гена А.Кернс-Смита (1985), согласно которой роль первичных репликаторов могли сыграть слоистые глины. Эти глины способны к ауторепликации слоев и обладают весьма сложной структурой, которой, по мнению Кернс-Смита, достаточно для мутаций и эволюции наподобие живых организмов. Глины эти постоянно кристаллизуются из слабых растворов кремниевой кислоты и гидратированных ионов металлов. Некоторые из таких кристаллов могли получить определенные преимущества благодаря захвату или синтезу на поверхности слоев органических соединений (см. выше гипотезу Берналла). Эти соединения могли играть роль катализаторов, увеличивающих скорость кристаллизации, изменять физические свойства глин, влиять на форму и размер кристаллов. Все это имело большое значение для репликации кристаллических глиняных генов. Предшественники РНК появились в достаточно развитых "глиняных организмах" и поначалу играли структурную роль. По мере совершенствования механизма саморепликации (или даже после становления системы синтеза белка) они покинули глиняный каркас и перешли к самостоятельному существованию, то есть на смену глиняному миру пришел мир РНК. Слабое место гипотезы "глиняного гена" состоит в том, что до сих пор в экспериментах с глиняными структурами не наблюдалось ничего похожего на эволюцию.
В последние десятилетия значительные изменения претерпевает и представление об условиях древней Земли. Дебатируется состав древней атмосферы, в которой, по мнению ряда исследователей, могли преобладать азот и углекислый газ, выделявшиеся вулканами. Примерно равновероятным признается возможность существования как восстановительной атмосферы, состоящей главным образом из СН4 + (N2 или NH3) + Н20, так и атмосферы с нейтральным окислительно-восстановительным потенциалом, в состав которой входят СО2 + N2 + Н20 (Chyba, Sagan, 1992). В принципе, и при том и ином раскладе химическая эволюция в том виде, в каком ее этапы были изложены выше, возможна, хотя второй вариант не слишком благоприятен для синтеза аминокислот. В соответствии с современными моделями происхождения планет Солнечной системы все более обоснованным выглядит предположение об изначально жарком климате на первых этапах эволюции Земли (Chang, 1999; Reysenbach et al., 1999). Поверхность планеты, кроме того, подвергалась в это время постоянным метеоритным бомбардировкам (Chyba, 1998), в том числе и гигантскими метеоритами диаметром более 10 км и кометами. При их падении на Землю выделялось огромное количество тепла, которого могло хватить, чтобы вода в океанах закипела. Все это делало условия существования на поверхности крайне нестабильными.
Не приходится удивляться, что поиски колыбели жизни переместились с поверхности Земли в глубь океана. Мощным толчком к этому послужило открытие в 70-х гг. морских гидротермальных источников на глубинах, куда не проникает солнечный свет. Вокруг них были обнаружены вполне процветающие экосистемы, роль первичных продуцентов в которых играли серобактерии, улавливающие соединения серы, выбрасываемые источниками. Учитывая, что ряд архебактерий, которые рассматриваются многими учеными как наиболее близко стоящие к предковым для всего живого формам жизни, являются экстремальными термофилами, гипотеза о возникновении жизни в районе гидротермален выглядит достаточно привлекательно. Впрочем, принимая в расчет представления о горячем климате древней Земли, протобионты в любом случае должны были быть приспособлены к существованию в условиях высоких температур.
Можно согласиться с мнением С.Чанга (Chang, 1999), который считает, что условия на границе раздела газовой, жидкой и твердой фаз, где имеются доступные источники энергии, в высокой степени благоприятны для этапов предбиоти-ческой эволюции и возникновения жизни, Этим критериям вполне соответствуют не только морские гидротермальные системы, но и зоны литорали, и перемешиваемый ветром поверхностный слой океана. Такие системы были широко представлены на древней Земле. Условия в них далеки от равновесных, причем имеется градиент свойств, поддерживаемый физическими и химическими потоками энергий. В рамках этих сред могли появляться многочисленные микромасштабные структуры с поверхностями разделов на основе разного рода аэрозолей, частичек вулканического пепла и пыли с суши, гидротермальных минералов, осаждающихся химических соединений, пузырьковидных структур органической и неорганической природы и т.п. Поскольку жизнь сама по себе возникла как некая пограничная структура (вспомним, к примеру, коацерваты), то способность окружающей среды образовывать, рассеивать и опять образовывать все новые и новые микроскопические структуры должна была послужить необходимой предпосылкой для зарождения жизни. Следует отметить, что в любом случае в рамках таких сред должны были существовать определенные процессы, снижающие риск вторичного разложения, захоронения или растворения компонентов, которым предстояло стать строительными блоками жизни.
Остается, наконец, упомянуть достаточно широкий спектр гипотез о заносе органических соединений и даже спор микроорганизмов (панспермия) из космоса. На возможность подобного сценария указывает обнаружение в некоторых метеоритах (углистые хондриты) органических соединений, включая аминокислоты (Хорганг, 1991). Органические вещества могут содержаться и в кометах. Имеются указания, что в некоторых углистых хондритах выявлены структуры, напоминающие споры бактерий и грибов. Все это не позволяет отбрасывать гипотезу о внеземном происхождении жизни, или, хотя бы, о прохождении определенных этапов химической эволюции вне нашей планеты. Хотя, если это и было действительно так, то нам придется искать другую планету, где имелись такие условия, которые способствовали бы начальным этапам эволюции жизни в той ее форме, которая столь хорошо приспособлена к существованию на Земле. По-видимому, проще представить, что все эти события имели когда-то место на нашей родной планете.
Глава 4 ОТ ПРОКАРИОТ К ЭУКАРИОТАМ
Где бы и как бы ни протекали первые этапы эволюции жизни, но примерно 3,5 млрд. лет тому назад на Земле уже существовали прокариотные организмы. Именно такой возраст имеют породы Варравуанской группы в Западной Австралии, в которых были найдены самые древние остатки прокариот, которых палеонтологи обозначают как микрофоссилии (Шопф, 1982; Маргелис, 1983, Schopf, 1999). Палеохимические исследования самых древних из известных на планете осадочных пород - формация Исуа в Гренландии - позволяют предполагать наличие микробной жизни значительно раньше, 3,8 млрд. лет назад (Крылов, 1985; Schidlowski, 1994). Кроме Австралии, микрофоссилии обнаружены в богатых углеродом кремнистых породах архейского времени в Южной Африке и Северной Америке, которые также относятся к древнейшим из известных неметаморфизированных пород. Палеонтологическая летопись позднего архея-протерозоя указывает на наличие в это время богатой микробной жизни, которая сформировала первую в истории Земли биосферу - прокариотную. Важнейшими ее компонентами были цианобактерии, следы жизнедеятельности которых обнаруживаются в виде карбонатных, иногда вторично окремненных слоистых отложений - строматолитов. Они формировались в результате накопления тонкого осадка, состоящего преимущественно из СаСО3, который захватывался фотосинтезирующими микроорганизмами. Живые строматолитовые маты и по сей день обнаруживаются кое-где на мелководьях тропических морей (например, залив Шарк-Бей на северо-западном берегу Австралии) и в солоноватоводных местообитаниях. В протерозое же, судя по палеонтологическим данным, строматолитовые ландшафты были широко распространены (Видаль, 1984; Розанов, Федонкин, 1994; Федонкин, 2000).
На протяжении архея и большей части протерозоя прокариоты безраздельно господствовали на Земле. В это время проходила их адаптивная иррадиация. Дискуссионным остается вопрос о способе питания первых живых организмов - протобионтов (Серавин, 1994). Долгое время считалось практически общепризнанным, что они должны были быть гете-ротрофами. Однако современные гетеротрофы для осуществления своих энергетических и синтетических процессов используют сложную систему ферментных реакций. Метаболизм же протобионтов несомненно был примитивен и для его осуществления они скорее всего нуждались в экзогенных источниках энергии, наиболее стабильным из которых могла быть только лучистая энергия Солнца. Поэтому вполне справедливым выглядит предположение Л.Н. Серавина (1994) о том, что протобионты были автогетеротрофами, точнее фотогетеротро-фами (Серавин, 1994). Это означает, что они использовали для построения своего тела органические молекулы, которые, скорее всего, в достаточно большом количестве имелись в первичных биотопах, какими бы они не были, а в качестве энергии - лучистую энергию. На основе примитивной фотогет-ротрофии у прокариот в дальнейшем формировался фотосинтез с участием хлорофиллов. На его раннее возникновение в истории жизни на Земле указывают палеохимические данные, согласно которым в осадочных породах возраста 3,5 млрд. лет обнаруживается повышенное содержание легкого изотопа 12С (известно, что фотосинтезирующие организмы легче усваивают 12СО2, чем 13СО2) (Schopf, 1999). Некоторые результаты исследований формации Исуа в Гренландии позволяют предполагать, что фотосинтетической активностью обладали уже организмы, жившие на планете 3,8 млрд. лет назад (Schidlowski, 1994). Гетеротрофия, по-видимому, развивалась вторично, возможно, после формирования озонового экрана, препятствующего доступу на поверхность планеты богатых энергией ультрафиолетовых лучей. Напомним, что само появление озонового экрана связано с накоплением в атмосфере кислорода, который имеет биогенное происхождение. Представления о первичной фотогетеротрофии протобионтов, хорошо согласующиеся с палеохимическими данными, затрудняют принятие гипотезы о возникновении и развитии жизни в глубинах океана, в районах с гидротермальной активностью (см. главу 3). Даже если признать подобный сценарий развития событий, то необходимо допустить, что уже на ранних этапах своей эволюции протобионты оказались на мелководьях или, по крайней мере, в пределах фотического слоя первичного океана, то есть там, где возможно существование фотогетеротрофов.
На уровне прокариот, очевидно, шла наработка различных биохимических механизмов усвоения и преобразования потребляемых клеткой химических соединений. Поводом для подобного предположения служат современные прокариоты, обладающие широчайшим разнообразием метаболических процессов, от факультативно анаэробного до, напротив, факультативно аэробного, с многочисленными переходными формами. Здесь мы сталкиваемся с разными типами брожений (спиртовое, молочнокислое, маслянокислое и др.), которые, несомненно, дали начало гликолизу эукариот. Именно у прокариот эволюируют цикл Кребса, дыхательная цепь и окислительное фосфорилирование, что, по сравнению с анаэробной утилизацией глюкозы, дает дополнительный выигрыш в 36 молекул АТФ в пересчете на одну молекулу глюкозы (Мусил и др., 1984). Развиваются различные способы получения энергии из химических соединений - хемосинтез. Наконец, как уже отмечалось чуть выше, формируются и разные варианты бактериального фотосинтеза на основе бактериохлорофиллов и, как завершающий этап биохимической эволюции - фотосинтез с участием хлорофиллов. Он распространен только среди цианобактерий и ничем принципиально не отличается от присущего фотосинтезирующим эукариотам.
По-видимому, именно цианобактерии сыграли неординарную роль в эволюции биосферы, поскольку с их активностью связывают накопление кислорода в атмосфере Земли. Строматолиты, образованные сообществами цианобактерий, становятся многочисленными в раннепротерозойских отложениях, образовавшихся примерно 2,3 млрд. лет назад. Выделявшийся стромалитообразующими цианобактериями кислород, по-видимому, вначале расходовался на окисление растворенного в первобытном океане железа, которое содержалось там в закисной (двухвалентной) форме. При окислении оно переходило в окисную (трехвалентную) форму и осаждалось в виде окислов, образовывавших слои окисных пород, так называемых полосчатых железняков (полосчатые железнорудные формации). Их образование происходило в относительно короткий (в геологическом масштабе) период в несколько сот миллионов лет, начавшийся немногим менее 2 млрд. лет тому назад. После окисления основной массы растворенного в океане железа кислород, выделяемый цианобактериями, стал в большом количестве поступать в атмосферу.
Судя по тому, что для всех эукариот (немногие исключения имеют вторичную природу - см. ниже) характерен аэробный метаболизм, их формирование происходило уже после накопления в атмосфере значительного количества свободного кислорода. Первые ископаемые эукариоты, так называемые акритархи, датируются возрастом примерно в 1,5 млрд. лет. Это небольшие, но крупные относительно бактерий, организмы (в среднем 10-500 мкм), одетые толстостенной оболочкой, часто орнаментированной, и имеющие сферическую или звездчатую форму (Видаль, 1984). Очевидно, это представители каких-то вымерших групп протистов. Таким образом, если на развитие прокариот, начиная с первых этапов химической эволюции, ушло около 500 млн. лет, то между первыми прокариотами и первыми эукариотами лежит неизмеримо больший отрезок времени, протяженностью примерно в 2 млрд. лет.
Ископаемая летопись не оставила нам следов начальных этапов формирования эукариот. Акритархи - это продукт достаточно длительной эволюции. Первые из них, найденные в формациях пород раннего рифея на Южном Урале (возраст 1,6-1,4 млрд. лет), по-видимому, близкородственны зеленым водорослям. Это уже весьма специализированные эукариоты, обладающие способностью к фотосинтезу. Поэтому, как и в отношении прокариот, мы можем только предполагать, каким путем возник этот новый уровень организации, с которым связана вся дальнейшая эволюция биосферы. В то же время задачу несколько облегчает то обстоятельство, что в данном случае нам достаточно хорошо известны начальный (бактериальный процит) и конечный (экариотная клетка - эуцит) продукты. Остается реконструировать промежуточные звенья, исходя из современных представлений о морфо-функциональ-ной организации прокариот и эукариот, и принимая в расчет те условия, которые существовали на нашей планете в середине-начале протерозоя. Возможные варианты таких реконст-пукиий, предлагавшиеся разными учеными, укладываются в памки двух основных концепций. Первая из них - это аутоге-нетическая концепция, согласно которой эукариотная клетка постепенно развивалась из прокариотного микроорганизма по мере совершенствования его организации (см. обзоры: Сера-вин, 1986а-г). Симбиотическая гипотеза, в наиболее ортодоксальной форме, предполагает, что формирование клетки эукариот происходило путем последовательных симбиозов нескольких прокариот (см. обзор: Маргелис, 1983). В последние годы наметилось объединение представлений об аутогенетическом и симбиогенетическом путях становления эукариотной клетки в рамках единой концепции, которая являет собой результат некоторого консенсуса между крайними точками зрения (Cavalier-Smyth, 1987; de Duve, 1996).
Предками эукариот, по-видимому, были гетеротрофные прокариотные микроорганизмы, которые жили в плотных бактериальных сообществах (весьма вероятно в поселениях циа-нобактерий - строматолитах, см. ниже) и питались, скорее всего, остатками отмерших организмов (de Duve, 1996). Переваривание пищи проходило внеклеточно, благодаря выделению наружу пищеварительных ферментов, что свойственно целому ряду современных микроорганизмов (рис. 6, а). Важнейшей предпосылкой для становления нового уровня организации послужила утрата жесткой клеточной стенки (рис. 6, б). Событие это легко представимое, учитывая, что среди современных микроорганизмов имеется такая группа, как микоплазмы, которые лишены клеточной стенки. Это чрезвычайно мелкие (0,2-0,3 мкм) симбиотические или паразитические бактерии, которые в силу отсутствия жесткого наружного каркаса, который представляет собой бактериальная клеточная стенка, способны менять форму тела. Утрата клеточной стенки сняла ограничения на рост микроорганизма (рис. 6, в). В условиях обилия пищи такие формы стали расти быстрее и достигать больших размеров, чем их родичи, окруженные плотной наружной оболочкой.
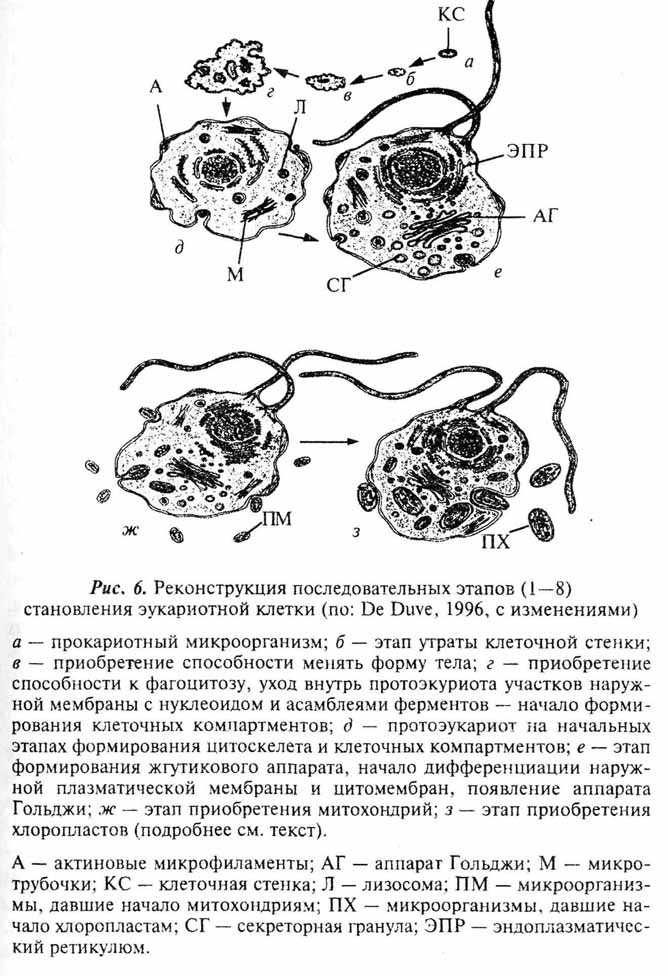
Увеличение размеров протоэукариот, по-видимому, привело к формированию питоскелета, который, кроме опорной функции, мог обеспечить и двигательную активность (Сера-вин, 1986 г) (рис. 6, г). Появление цитоскелета, конечно, еще не в той полной и совершенной форме, в которой он обнаруживается у современных эукариот, послужило следующей важной посылкой на пути становления нового уровня организации живого. У имевших его организмов участки поверхностной мембраны могли не только инвагинировать, но и отрываться в виде замкнутых пузырьков. На этой основе развились характерные для клеток современных эукариот механизмы пино- и фагоцитоза. Это кардинальным образом изменило способности организма по части использовании источников питания. Появилась возможность заглатывать крупные частицы пищи и даже другие микроорганизмы, то есть наметился переход к хищничеству. Это должно было существенно повысить конкурентоспособность проэукариот, поскольку они сформировали новые экологические ниши - крупных (в масштабах микробных сообществ, естественно), активно перемещающихся хищников и детритофагов.
Становление этих проэукариот сопровождалось и иными не менее значимыми перестройками б их морфо-функцио-нальной организации. Участие наружной мембраны в обеспечении движения, фаго- и пиноцитоза сделало весьма неудобным расположение на ее внутренней поверхности или впячи-ваниях мезосомального типа ассамблей ферментов, обеспечивающих те или иные этапы метаболизма. Такие участки стали отшнуровываться от поверхностной мембраны в виде вакуолей разного размера и формы, которые дачи начало лизосомам, ЭПР, комплексу Гольджи и другим органеллам (рис. 6, д). В результате в протоклетке происходило формирование различных клеточных компартментов - характерная черта цитоплазмы эукариотного типа. Параллельно изменялась и наружная мембрана. Перемещение внутрь клетки ферментных систем изменило химизм поверхностной мембраны и повлекло за собой дальнейшие биохимические преобразования (Серавин, 1986 г). В составе мембраны появились стеролы (в первую очередь, холистерол), что сделало ее лабильной, повысило способность к везикулизации (см. выше).
Таким образом, постепенно формировались цитоплазма и мембраны эукариотного типа. Это способствовало усовершенствованию обмена веществ и механизмов распределения питательных соединений внутри организма, что, в свою очередь, вело к еще большему увеличению размеров проэукариот. По мере функциональной дифференциации органелл их мембраны по своему белковому составу стали отличаться от наружной, что в конечном итоге привело к формированию присущей современной эукариотной клетке системе внутренних ци-томембран и наружной плазматической мембране.
Постепенно размеры проэукариот достигли некоторого критического уровня, когда регуляция обменных процессов небольшим нуклеоидом стала затруднительной. К этому времени нуклеоид, скорее всего, уже переместился внутрь цитоплазмы. Причины и механизм этого перемещения были, по всей видимости, те же, что и при отшнуровывании несущих ферменты участков поверхностной мембраны, которые дали начало ряду клеточных органелл. Можно предположить, что увеличение объема цитоплазмы у проэукариот привело к увеличению информационной емкости их нуклеоида. Происходило это, скорее всего, в результате умножения числа копий той единственной кольцевой молекулы ДНК, которая изначально входила в его состав (Серавин, 1986 г.). Речь, таким образом, идет об умножении числа геномов и появлении полигеномных нуклеоидов, состоявших из двух и более одинаковых кольцевых молекул ДНК. Такого рода полигеномные нуклеоиды обнаруживаются у некоторых современных цианобактерий (Громов, 1983). Коль скоро появилось несколько копий одной молекулы ДНК, то в них неизбежно стали накапливаться разные мутации, что постепенно привело к формированию существенных генетических различий между входящими в состав полигеномного нуклеоида молекулами ДНК. Можно предположить, что эти различающиеся последовательностями нукле-отидов ДНК дали начало хромосомам. В ходе их эволюции имел место переход от кольцевой к линейной структуре ДНК, установление связи с белками, в первую очередь гистонами, и, в конце концов, формирование хромосом в том виде, в каком они присущи ядерному аппарату современных эукариот (Серавин, 1986 г.). Возможно, первичные хромосомы свободно лежали в цитоплазме, а ядерная оболочка представляет собой более позднее эволюционное приобретение. Процесс ее формирования мог заключаться в особой дифференциации цистерн эндоплазматического ретикулюма.
На одном из ранних этапов эволюции проэукариот произошло и формирование жгутика (рис. 6,е). Его появление связано, по всей видимости, с псевдоподиеподобными выростами цитоплазмы, которые на начальных этапах эволюции проэукариот обеспечивали перемещение по субстрату и/или захват пищи. Такого рода выросты характерны для многих современных протистов (щупальца, ретикулоподии, аксоподии и др.) (Серавин, 1985). В них всегда имеются тем или иным способом упакованные микротрубочки, на основе которых, по всей видимости, формировался сократительный аппарат ундулиподии (внешняя, рабочая часть жгутика, способная совершать ритмические движения). Параллельно идет и образование базальной гранулы жгутика или кинетосомы, которая функционирует как центр сборки сократимых систем ундулиподии. На раннее появление жгутика (реснички) в ходе эволюции эукариот указывает то обстоятельство, что все современные организмы обладают им в той или иной форме на какой-либо стадии жизненного цикла. Современные безжгутиковые формы, скорее всего, имеют вторичное происхождение, хотя нельзя окончательно исключить их раннее ответвления от общего филогенетического ствола эукариот на дожгутиковой стадии. Необходимо отметить, что от кинетосомы жгутика (реснички) берут начало центриоли, которые аналогичны им по строению и служат центром образования микротрубочек митотического веретена в ходе клеточного деления. Известно, что кинетосомы некоторых протистов могут брать на себя функцию центриолей, а последние, наоборот, способны формировать несовершенные реснички (Серавин, 1985).
Все описанные выше события вполне укладываются в сценарий гипотезы об аутогенном происхождении клетки эукарот (Серавин, 1986). Объяснение же с этих позиций становления таких органелл, как митохондрии и хлоропласты, наталкивается на целый ряд серьезных трудностей. И те, и другие окружены двойной мембраной (у хлоропластов некоторых протистов число мембран более двух - см. главу 7). Причем, если наружная вполне соответствует по своей организации эукариотной, то внутренняя больше напоминает поверхностную мембрану бактерий. Обе рассматриваемые органеллы имеют собственные рибосомы, сходные по структуре с бактериальными (70S), свою кольцевую ДНК, и обладают способностью к делению вне зависимости от стадии клеточного цикла. То, что митохондрии и хлоропласты выделяются в ряду остальных клеточных органелл, было подмечено давно и уже более ста лет тому назад высказывались предположения об их симбиотическом происхождении (рис. 6,ж,з). По мере развития методической базы современной клеточной биологии это предположение, изначачьно воспринимавшееся "в штыки", находит все больше подтверждений (см. обзоры: Lang et al., 1999; Turner et al., 1999). Относительно симбиотического происхождения хлоропластов практически все сомнения к настоящему времени рассеялись. Возможные пути становления этих органелл в разных группах фотосинтезирующих эукариот мы рассмотрим отдельно в главе 7. Недавние молекулярно-генетические исследования митохондриальных генов протистов значительно укрепили и гипотезу о происхождении митохондрий от внутриклеточных симбионтов бактериальной природы (Gray et al., 1999; Lang el al., 1999). Более того, ряд специфичных особенностей в организации митохондриальных генов свидетельствует, что процесс приобретения этой органеллы в ходе эволюции эукариот произошел единожды. Иными словами, митохондрии у всех ныне живущих эукариот ведут свое начало от общего бактериального предка.
Полезность подобного приобретения для проэукариотного организма, который не обладал циклом Кребса, дыхательной цепью и окислительным фосфорилированием, несомненна. Достаточно вспомнить, что в результате цикла трикарбоновых кислот на каждые две молекулы пирувата (а следовательно, на каждую молекулу глюкозы, вступающую в цикл) образуются в конечном итоге 34 добавочные молекулы АТФ. Очевидно, что эндосимбиоз мог возникнуть только после приобретения про-эукариотами способности к фагоцитозу. Иного пути проникновения в клетку, чем внутри фагоцитарной вакуоли, для инородных частиц, в том числе и пиши, не существует. На принципиальную возможность же эндосимбиоза эукариот с прокариотами указывают многочисленные современные примеры (см. главу 7).
Видимо, какие-то фагоцитированные проэукариотами бактерии, обладающие системами аэробного расщепления са-харов, не переваривались, а оставались в цитоплазме как внутриклеточные симбионты (рис. 6,ж). Они были окружены двойной мембраной; наружная представляла собой мембрану фагоцитарной, или, точнее эндосибиоцитозной, вакуоли (эукариотная организация), а внутренняя - наружную мембрану микроорганизма-эндосимбионта (прокариотная организация). В эндосимбионте поначалу сохранялись все присущие прокариотам структуры, в том числе нуклеоид, рибосомы, ферментные системы. По мере углубления эндосимбиотических отношений произошло упрощение организации эндосимбионта-промито-хондрии, который из самостоятельного микроорганизма эво-люировал в клеточную органеллу. Это неизбежно сопровождалось сокращением числа генов в составе митохондриальной ДНК и привело к тому, что часть митохондриальных белков у современных эукариот кодируется ядерной ДНК.
Напомним, что речь идет об очень древнем эндосимбиозе, возникшем на заре становления эукариот в далеком протерозое. Митохондрии были присущи общему предку всех современных эукариот, поскольку все они таковыми обладают. Сравнительно недавно было установлено, что митохондриаль-ные гены (ядерные гены, кодирующие некоторые митохонд-риальные белки) имеются в геномах ряда паразитических простейших (микроспоридии, трихомонады и дипломонады - подробнее см. главу 7), у которых митохондрии отсутствуют (Muller, 1997; Hasegawa, Hashimoto, 1999). Это говорит об их уже вторичной утрате, вероятно, в связи с паразитическим образом жизни в анаэробных условиях. Таким образом, новые данные опровергли еще недавно бытовавшие в литературе суждения о наличии среди современных эукариот реликтов домитохондриальной эры и заставили пересмотреть некоторые взгляды на филогенетические взаимоотношения между представителями разных групп протистов (см. главу 7).
Помимо несомненного энергетического выигрыша от эндосимбиоза проэукариот с митохондриеподобными бактериями установлению такого рода отношений могло способствовать и другое не менее, если не более, важное обстоятельство. Речь идет о том, что Де Дюв (de Duve, 1996) назвал "кислородным холокостом". Действительно, накопление кислорода в атмосфере протерозоя связано с фотосинтетической активностью микроорганизмов и, прежде всего, цианобактерий, широко к тому времени распространившихся. Интенсивность этого процесса нарастала примерно со времени 2 млрд. лет тому назад, пока около 1,5 млрд лет тому назад не был достигнут некоторый относительно стабильный уровень (примерно 1% от современного уровня содержания кислорода в атмосфере - Нолл, 1991). До появления в атмосфере кислорода все существовавшие в тогдашней биосфере формы жизни были адаптированы к анаэробным условиям. Очевидно, что, как и современные анаэробы, они были крайне чувствительны к присутствию кислорода. Дело в том, что при попадании в клетку кислорода образуется ряд токсичных соединений, таких как пероксидные ионы, гидроксильные радикалы и перекись водорода. Поэтому можно предположить, что многие древние анаэробы, которые не смогли адаптироваться к новым условиям, вымерли в результате "кислородного холокоста". Анаэробом был и наш передковый эукариотный организм, для которого приобретение эндосимбионта, способного утилизировать кислород и переводить его в безвредную воду, дало шанс на выживание. Не говоря уж об энергетическом выигрыше, речь о котором шла выше.
Таким образом, современная гипотеза эволюции эукари-отной клетки в своей основе остается все же аутогенетичес-кой. Она постулирует постепенное перерастание пронитной, пользуясь терминологией Л.Н.Серавина, организации прокариот в тот сложный морфо-функпиональный ансамбль, который являет собой эукариотная клетка. На определенном этапе этой эволюции, когда предковой формой уже был достигнут, по сути дела, эукариотный характер организации цитоплазмы, появилась возможность использовать механизм эндосимбиоза. Такая возможность и была реализована, что, несомненно, в значительной степени способствовало повышению конкурентоспособности проэукариот и их широкому распространению в биотопах протерозоя.
Глава 5
"ВОЦАРЕНИЕ" ЭУКАРИОТ
Началу экспансии эукариот в немалой степени способствовали изменения среды обитания, происходившие на протяжении протерозоя (примерно 2,6-0,6 млрд. лет тому назад) (Федонкин, 1987, 2000; Розанов, Федонкин, 1994). Постепенно снижались вулканическая и гидротермическая активности, огромные запасы жизненно важных химических соединений были изъяты из биогеохимического круговорота и захоронены в виде рудных залежей осадочного происхождения. Таким образом, значительно уменьшился объем геохимических ресурсов энергии, к использованию которых прокариоты приспособились на протяжении всей своей предшествующей эволюции. Имело место и существенное понижение температуры, которая в ряде местообитаний могла переходить уровень, критический для древних прокариот, чье формирование проходило в гораздо более горячих условиях (см. выше). Интересно, что значительный вклад в охлаждение окружающей среды внесли сами микроорганизмы. Роль пускового механизма сыграло, по предположению А.Ю.Розанова и М.А.Федонкина (1994), формирование обширных континентальных плит. С этим процессом связано увеличение площади мелководий и возникновение явления апвеллинга. Последний обеспечивает вынос на поверхность холодных глубинных вод, обогащенных химическими веществами (главным образом, фосфатами и нитратами), которые необходимы для фогосинтезирующих организмов. В районах современных апвеллингов наблюдаются самые высокие показатели первичной продукции в результате массового размножения диатомовых водорослей и других представителей фитопланктона. Создаваемая этими водорослями биомасса поддерживает существование богатейших по обилию и разнообразию экосистем. Достаточно вспомнить знаменитый Перуанский апвеллинг, благодаря которому прибрежье Перу стало одним из наиболее продуктивных районов современного Мирового океана (Айдилл, 1981).
Наличие обширных мелководий и необходимых химических соединений, несомненно, способствовали расцвету фотосинтезирующих микроорганизмов и в первую очередь цианобактерий. Свидетельство тому - многочисленность протерозойских строматолитов в палеонтологической летописи. Рост биомассы цианобактерий сопровождался изъятием огромных количеств углекислоты из воды и атмосферы и биогенным карбонатоосаждением. Поскольку углекислый газ является одним из важнейших компонентов парниковых газов, то его интенсивное изъятие привело к снижению парникового эффекта (К парниковым газам относятся углекислый газ, метан, окислы азота и серы и ряд других, которые пропускают солнечную радиацию, но задерживают исходящее от поверхности Земли длинноволновое тепловое излучение. В результате создается "эффект парника". Если бы в составе современной атмосферы парниковые газы отсутствовали, то поверхность планеты охладилась бы в среднем до температуры минус 18 °С). Кроме того, выделявшийся в процессе жизнедеятельности цианобактерий кислород окислял и другой парниковый газ - метан. Все это вело к снижению температуры окружающей среды, которое сопровождалось повышением концентрации кислорода в атмосфере. Последний, как уже отмечалось выше, оказывал угнетающее воздействие на многих прокариот, в том числе и на "породивших" его цианобактерий (Крылов, 1985). Таким образом, сами микроорганизмы в значительной степени способствовали глобальному преобразованию окружающей среды таким образом, что многие их группы вымирали, другие вынуждены были ограничить свое распространение оставшимися местообитаниями с анаэробными условиями существования, иные же приспосабливались к жизни в кислородном окружении.
Как уже отмечалось выше, становление эукариот должно было происходить ранее 1,5 млрд. лет назад (время, которым датируются самые ранние из обнаруженных акритархов). Это период расцвета строматолитообразующих цианобактерий и при относительно еще невысоком содержании кислорода в атмосфере (напомним, что первоначально выделяемый циано-бактериями кислород расходовался преимущественно на окисление железа и других металлов, что привело к образованию осадочных руд). В то же время локально высокие концентрании кислорода существовали даже в архее в тонком (живом) поверхностном слое строматолитов (цианобактериальные маты), особенно широко распространившихся в протерозое (Розанов, федонкин, 1994). Именно здесь, по предположению А.Ю. Розанова и М.А. Федонкина (1994), и проходило формирование первых эукариот, которые первоначально утилизировали продукты жизнедеятельности цианобактерий. С матами цианобактерий связаны весьма разнообразные микробные сообщества, которые могли послужить источниками микроорганизмов, входивших в эндосимбиоз с проэукариотами. Некоторые из сформировавшихся эукариот стали покидать свой первичный биотоп и переходить к существованию в обогащенной кислородом морской воде. Недаром большинство акритархов представляют собой планктонные организмы, пик разнообразия которых приходится на период 900-800 млн. лет назад (Видаль, 1984; Розанов, Федонкин, 1994). Появившиеся эука-риотные водоросли успешно конкурировали с цианобактерия-ми за ресурсы и местообитания, они активно расселялись, увеличивали свою численность и тем самым способствовали еще большему обогащению гидросферы и атмосферы кислородом и, как следствие, снижению парникового эффекта.
Все вместе взятое постепенно привело к закату господства прокариот и к глобальному изменению биосферы, которое И.Н.Крылов (1985) образно именует первой глобальной экологической катастрофой. Пик разнообразия и численности строматолитовых сообществ приходится на период 1 млрд. лет назад, вслед за чем следует резкий спад. В немалой степени тому, по всей видимости, способствовала и длительная ледниковая эпоха, наступившая около 850 млн. лет назад. С этого времени и вплоть до начала кембрия биосфера испытала серию перестроек, во многом связанных с серией мощных покровных оледенений. Причины глубокого и длительного похолодания на планете пока не вполне понятны. Однако весьма вероятной представляется их связь с уменьшением парникового эффекта, вызванного как биогенными причинами, речь о которых шла выше, так и выводом из круговорота значительной части биогенного углерода в результате его захоронения на обширных и стабильных континентах (Нолл. 1992; Федонкин, 2000). В древние ледниковые периоды, длительность которых составляла от 4 до 30 млн. лет, льды достигали уровня моря даже на континентах, расположенных вблизи экватора (Чумаков, 1992). Угнетению подверглись строматолитовые сообщества морских мелководий, вымерли практически все планктонные акритархи размером более 600 мкм. В то же время общее похолодание в немалой степени способствовало "воцарению" эукариот, на что указывают многие особенности вендской фауны.
Именно ко времени позднего протерозоя, к венду (от 700-650 до 550-590 млн. лет назад), относятся первые находки крупных многоклеточных животных. Обнаружены они были еще в 40-х годах, но переосмысление этих находок, вызвавшее сенсацию в научном мире, произошло относительно недавно, начиная с 60-х гг. До этого времени общепризнанным считалось, что первые многоклеточные появились в кембрии, то есть около 550 млн. лет назад. Открытие вендской фауны и ее исследование заставило во многом пересмотреть устоявшиеся взгляды на эволюцию многоклеточных эукариот. Вендские организмы впервые были обнаружены в Австралии, в местности Эдикар (отсюда берет начало название второй половины вендского периода - эдикарская эпоха и часто встречающееся в литературе наименование "эдикарская фауна"), а затем последовали многочисленные находки, сделанные на Зимнем Берегу Белого моря (Федонкин, 1981, 1987; Соколов, 1984). Характернейшей особенностью вендских животных является отсутствие минерализованного скелета ("бесскелетная фауна венда") и крупные, даже гигантские размеры многих представителей. Среди вендских животных резко преобладали кишечнополостные, в числе которых огромные медузы с колоколом, достигающим метрового диаметра, сидячие медузы, одиночные и колониальные полипы. Кроме кишечнополостных, достаточно широко представлены биллатерально-сим-метричные животные, в том числе и сегментированные. Это плоские и кольчатые черви, примитивные членистоногие и моллюски (Федонкин, 1981, 1998). Помимо представителей филогенетических линий, имевших продолжение в кембрии и затем среди современных животных, встречаются и многоклеточные животные, принадлежащие к группам, полностью вымершим в ходе биосферных перестроек вендского периода.
О холодноводности вендской фауны говорит уже тот факт, что ископаемые останки вендских животных обнаруживаются в породах, формирование которых могло происходить только в условиях умеренного или даже холодного климата (Федонкин, 2000). Другим указанием служит отсутствие минерального (известкового) скелета, построенного из карбонатов. Дело в том, что растворимость карбонатов значительно выше в холодных водах, и поэтому энергетические затраты на постройку состоящего из них скелета в холодных водах существенно выше, чем в теплых. Этот феномен объясняет, почему скелет животных, например, раковины моллюсков, обитающих в тропиках, значительно массивнее, чем у близкородственных холодноводных форм. Наконец, судя по палеонтологическим данным, вендские трофические цепи были коротки, что так же характерно для современных холодноводных бассейнов (например, фитопланктон - зоопланктон - кит). Таким образом, предположение о том, что вендская фауна была приурочена к холодным водам, выглядит весьма обоснованным.
В начале венда имело место одно из крупнейших покровных оледенений в истории Земли - варангерское оледенение (650-700 млн. лет назад). Оно привело к охлаждению мирового океана и сокращению, таким образом, местообитаний, пригодных для развития сообществ микроорганизмов. Последние локализовались в теплых мелководных бассейнах и полностью контролировали моря протерозоя вплоть до венда. Надо отметить, что многие современные цианобактерии выделяют вещества (алкалоиды, пептиды и птеридины), ядовитые для эукариот (Саут, Уиттик, 1990). Это одна из причин, почему "цветение" воды, вызываемое цианобактериями, является настоящим бедствием для водоемов. Существует определенный антагонизм между цианобактериями и беспозвоночными животными, подтверждением чему может служить крайняя бедность эукариотной жизни в районах современных поселений стро-матолитообразующих цианобактерии. Видимо, то же самое имело место и в условиях тепловодных бассейнов протерозоя с богатым и разнообразным микробным населением, основу которого составляли цианобактерии. Поэтому появление обширных холодноводных акваторий дало шанс эукариотам эволюировать в условиях отсутствия конкуренции со стороны устоявшихся микробных сообществ. И они этим шансом воспользовались.
Следует отметить еще одну важную особенность холодных вод - их значительную насыщенность кислородом по сравнению с водой в более прогреваемых бассейнах. Вендские животные, эволюция которых проходила в холодноводных морях, несомненно, были адаптированы к условиям существования в воде, относительно богатой кислородом. Возможно, это было одной из причин, позволивших вендским эукариотам значительно увеличить размеры тела. Дело в том, что существует определенный кислородный барьер, составляющий не менее 6-10% от концентрации кислорода в современной атмосфере, ниже которого развитие крупных аэробных организмов невозможно (Нолл, 1991). Экспансия уже значительно продвинутых в эволюционном плане вендских животных в обедненные кислородом теплые воды стала возможной только после резкого возрастания в атмосфере свободного кислорода. Это событие произошло в самом конце протерозоя (Нолл, 1991).
Проникновение в теплые моря сняло ограничения на формирование минерального скелета. Именно этим может объясняться массовое развитие скелетных форм в кембрии. Находит, таким образом, свое объяснение и проблема "кембрийского взрыва" эволюции, которая волновала биологов со времен Ч. Дарвина. Действительно, долгое время палеонтологическая летопись начиналась только с кембрия, когда уже были представлены все основные типы животных. Вставал вопрос, когда же они успели сформироваться? Что послужило причиной "кембрийского взрыва", протяженность которого оценивается всего в 5-10 млн. лет, то есть одно мгновение по масштабам геологического времени? Описанные выше представления об условиях эволюции эукариот в конце протерозоя позволяют ответить на эти вопросы. Видимо, именно в кембрии животные широко распространились на прогреваемых мелководьях, и многие их представители обзавелись минерализованным скелетом. Шансы же на сохранение в ископаемой летописи скелетных форм неизмеримо больше, и таким образом "кембрийский взрыв" представляет собой скорее "взрыв изобилия окаменелостей", чем эволюционное событие (Bell, 1997; Федонкин, 2000).
Итак, "кембрийскому взрыву" предшествовала эволюция бесскелетных вендских животных. Но и последние не могли появиться на пустом месте. Какие-то предковые многоклеточные животные несомненно существовали в довендское время. Очевидно, они были мелки и не обладали развитыми скелетными элементами органической или минеральной природы. В силу этого шансов найти следы этого раннего этапа эволюции многоклеточных в ископаемой летописи немного. Однако косвенным подтверждением существования довендских многоклеточных животных служат молекулярно-генетические данные, полученные с использованием метода молекулярных часов (см. выше). В масштабном исследовании, выполненном Реем, Левинтоном и Шапиро (Wray et al., 1996), было проведено сравнение последовательностей семи генов, не связанных функционально (ядерные и митохондриальные гены), и 18S рРНК у значительного числа представителей разных систематических групп животных. Обнаружилось, что расхождение филогенетических ветвей первичноротых (в анализ были вовлечены моллюски, кольчатые черви и членистоногие) и вторичноротых животных произошло раньше, чем разделение вторичноротых на иглокожих и хордовых. Это вполне соответствует представлениям зоологов. Сенсационной оказалась датировка этих событий: 1,2 млрд. лет назад для первого и 1,0 млрд. лет для второго. То есть задолго до начала венда, не говоря уж о кембрии. Конечно, датировки с использованием метода молекулярных часов достаточно спорны, о чем мы уже писали, и вокруг результатов анализа Рея, Левинтона и Шапиро ведутся жаркие споры (см., например, Bell, 1997). Однако представления о том, что ранняя эволюция многоклеточных проходила в довендское время, вряд ли подвергнутся сомнению.
Глава 6
"УРОВНЕВЫЙ" ПОДХОД К ПОСТРОЕНИЮ МЕГАСИСТЕМЫ
В трех предыдущих главах мы попытались обозначить основные вехи на пути эволюции двух фундаментальных уровней организации живых систем: прокариот и эукариот. Прокариоты, как мы видели, достаточно естественно подразделяются на две большие группы: архебактерии и эубактерии. Сложнее обстоит дело с попыткой выделения крупных градаций внутри эукариот, учитывая, что доминировавшее долгое время подразделение на растения и животные, как было показано ранее, не может считаться адекватным накопленному массиву данных. Один из возможных вариантов решения задачи состоит в использовании "уровневого" подхода, методология которого была описана в главе 2.
Не приходится сомневаться, что первые эукариотные организмы были одноклеточными. Причем одноклеточными в том понимании, которое применимо к современным одноклеточным протистам. Речь идет о клетке как организме, который, в отличие от клеток, входящих в состав тканей многоклеточных животных, обладает набором морфо-функдиональных адаптации, позволяющих ему самостоятельно существовать в той или иной окружающей среде (Полянский, 1971). Так, у современных одноклеточных протистов обнаруживаются наборы органелл и особенности организации цитоскелета, которые не встречаются у клеток многоклеточных с тканевой организацией (Серавин, 1991).
На определенном этапе эволюции одноклеточных древних эукариот неизбежно встал вопрос об увеличении размеров тела и совершенствовании морфо-функциональной организации. Это значительно повышало шансы в конкурентной борьбе, прежде всего в силу возможности использовать новые источники пищи (более крупные органические частицы, других протистов и т.п.). Однако, как известно, клетка не может беспредельно увеличивать свои размеры, поскольку ядро способно контролировать только ограниченный объем цитоплазмы. Попытки выхода из этой казалось бы тупиковой ситуации можно проследить при анализе современных эукариот.
Сразу приходит на ум вариант увеличения числа ядер. Он реализуется у целого ряда протистов. Например, у комменсальных жгутиконосцев-опалин число ядер может достигать 100 и более, размеры же тела доходят до 1 мм, то есть превышают значения, характерные для многих многоклеточных, таких как некоторые плоские черви, коловратки и т.п. Умножение числа ядер характерно и для представителей одной из крупнейших групп протистов - фораминифер. У ряда представителей этой группы обнаружена дифференциация ядер на вегетативные и генеративные. Первые берут на себя контроль за клеточным метаболизмом, а вторые дают начало новому поколению. При этом специализация в сторону вегетативного ядра необратима, генеративное же ядро, в случае необходимости, может дать начало вегетативному. Последнее отличается большими размерами и количеством РНК, в то время как количество ДНК не увеличивается (Михалевич, 2000). Форами-ниферы считаются гигантами среди протистов. Размеры некоторых их современных и вымерших представителей достигают 12-16 см (наибольшую известность в этом плане получили вымершие формаиниферы - нуммулиты) (Михалевич, 2000).
Широкое распространение получили и разного рода многоядерные плазмодии, которые образуются в результате не доведенного до конца деления или слияния нескольких одноклеточных особей. Возможно и сочетание двух этих процессов, что наблюдается у некоторых миксомицетов. Первичный плазмодий возникает при слиянии нескольких особей, вслед за чем следует деление ядер, не сопровождающееся разделением цитоплазмы. Такие плазмодии могут иметь очень большие размеры. Так у миксомицета Fuligo septica они достигают 1,5 м в длину (Серавин, 1991, 2000).
Во многом сходны с плазмодиями и талломы некоторых водорослей, организацию которых принято именовать сифо-нальной (Саут, Уиттик, 1990). Этот тип организации независимо возник среди зеленых и желто-зеленых водорослей. Пожалуй, наиболее сложно устроены сифоновые водоросли (зеленые), тело ряда представителей которых (каулерповые) сильно расчленено и может имитировать многоклеточный растительный организм. По поверхности грунта стелются цилиндрические части слоевища, от которых вниз и вверх отходят отростки. Нижние, ризоиды, обеспечивают плотное прикрепление слоевища к любому субстрату, а отходящие вверх ветвятся на концах и функционально соответствуют листьям растений, поскольку именно в них сосредоточены хлоропласты. В целом же слоевище лишено внутренних перегородок и представляет собой, по сути дела, многоядерный плазмодий. Если же перегородки имеются, то все равно каждый отдельный сегмент всегда многоядерный (Виноградова, 1977).
Талломы красных водорослей, также достигающие весьма сложной организации, состоят из многих клеток. Однако между этими клетками имеются поры, благодаря натичию которых цитоплазма всех составляющих таллом клеток оказывается связанной в единое целое, что напоминает организацию син-цитиальных тканей животных (Серавин, 1991).
Еще один возможный вариант увеличения размеров одноклеточного организма лежит на пути интенсификации работы ядер. Начальные этапы этого процесса прослеживаются у тех фораминифер, ядра которых подразделяются на одно или несколько вегетативных и многочисленные генеративные. Вегетативное ядро крупное, иногда полиплоидное, и обеспечивает все жизненные отправления организма, в то время как генеративные участвуют только в процессах воспроизводства. Гораздо в большей степени дифференцированы ядра большинства инфузорий, которых нередко рассматривают как вершину эволюции одноклеточных организмов. Так же как и для фораминифер, им присущ ядерный дуализм, то есть подразделение ядер на вегетативное (макронуклеус) и генеративное (микронуклеус) (Райков, 1978). Плоидность макронуклеуса достигает нескольких тысяч, причем у высших инфузорий (брюхорес-ничные) наблюдаются достаточно сложные генетические перестройки. В процессе роста макронуклеуса вначале происходит умножение числа хромосом, затем они распадаются и происходит деградация части ДНК. Остаются, по-видимому, только те гены вместе с контролирующими их экспрессию участками, которые необходимы для нормального обеспечения вегетативных функций инфузории. Эти участки ДНК затем еще неоднократно реплицируются. В результате в зрелом макронуклеусе оказывается огромное количество копий структурных генов. В то же время микронуклеус всегда диплоиден и не испытывает никаких генетических реорганизаций. В нем в неприкосновенном виде консервируется характерный для данной особи геном.
Макронуклеус не может функционировать беспредельно. По мере старения инфузории неизбежно накапливаются сбои в считывании и передаче генетической информации, что проявляется в угнетении активности особи. Для предотвращения этих негативных явлений служит половой процесс, в начале которого микронуклеус претерпевает мейоз. Из образовавшихся в его ходе четырех гаплоидных ядер два резорбируются, а два оставшихся либо сливаются друг с другом (аутогамия), либо же, в случае настоящей конъюгации (тесное соединение друг с другом двух входящих в половой процесс инфузорий, в ходе которого между ними формируется цитоплазматический мостик), конъюгирующие особи обмениваются одним из гаплоидных ядер, вслед за чем в каждой из них происходит слияние "чужого" ядра с материнским. В любом случае образуется диплоидное ядро, которое делится и восстанавливает характерное для данного вида число макро- и микронуклеусов. Старый же макронуклеус дегенерирует.
Описанные особенности строения макронуклеусов позволяют инфузориям многократно повысить интенсивность метаболических процессов. Не удивительно, что размеры этих протистов весьма велики и у некоторых видов могут превышать 1 мм. Что касается сложности организации, то она до некоторой степени сравнима с таковой у многоклеточных животных. Так у паразитирующих в рубце жвачных инфузорий энтодини-умов на переднем конце имеется сложно устроенный ресничный коловращательный аппарат. Захваченная им пища поступает в специализированный участок цитоплазмы - клеточный рот, который ведет в "глотку" со сложно дифференцированным цитоскелетом. Имеется и "задняя кишка" - участок цитоплазмы на заднем конце тела, где происходит выброс наружу непереваренных остатков из пищеварительных вакуолей. И все это на клеточном уровне.
Наконец, еще один возможный механизм увеличения размеров - это умножение числа входящих в состав организма клеток. Здесь имеется множество вариантов. Один из них реализуется у некоторых амебоидных организмов, например, у диктиостилид. Отдельные амебки на определенной стадии жизненного цикла сползаются вместе и образуют клеточную массу, которая функционирует как самостоятельный организм - псевдоплазмодий. В отличие от настоящего плазмодия, который формируется в результате не доведенных до конца делений, в этом случае имеет место агрегация клеток, имеющих разное происхождение. Псевдоплазмодий диктиостилид способен перемещаться по субстрату и по прошествии некоторого времени формирует плодовое тело. Часть клеток образуют его подошву, другие - стебелек, на конце которого имеется расширение, формируемое клетками, которые дифференцируются в споры. Образование псевдоплазмодиев характерно также для акразиевых и некоторых других протистов, но в целом агрегации не получили широкого распространения.
По-видимому, наиболее продуктивным в эволюционном плане оказалось формирование многоклеточных организмов в результате нерасхождения клеток, образовавшихся в ходе деления. При этом отдельные клетки, происходящие от одной материнской, удерживаются друг с другом механически при помощи слизи и т.п. На начальном этапе этого процесса, по всей видимости, появлялись нестабильные объединения только что разделившихся клеток. Современные примеры такого рода дают так называемые временные колонии, характерные, в частности, для широко распространенных представителей рода СегаПит (динофитовые). Разделившиеся клетки этих ди-нофитовых некоторое время остаются связанными друг с другом и образуют цепочки. От такого рода временных колоний ведут два пути. Один - это формирование разнообразных многоклеточных слоевищ, а второй - совершенствование колониальной организации.
Первый путь избрали многие низшие растительные организмы. Это привело к появлению того многообразия нитчатых, паренхимных и иных типов талломов, которое характерно для багрянок, золотистых, желто-зеленых, зеленых, бурых и ряда других водорослей. Особого развития паренхимная организация достигла у некоторых представителей бурых, которые вплотную подошли к формированию настоящих тканей (Саут, Уиттик, 1990). Так у высокоспециализированных ламинарий и фукусов таллом отчетливо подразделяется на пластинку, ножку и ризоиды. Развиты коровые и сердцевинные ткани, поверхностные слои клеток продолжают делиться на протяжении всего периода роста таллома и получили название мерисгодер-мы. Транспорт метаболитов может осуществляться через специальные трубчатые структуры, которые аналогичны флоэме высших растений.
Колониальность особо широкое распространение получила у зеленых водорослей, а среди них у вольвоксовых, в рамках которых прослеживаются практически все этапы формирования колоний. В начале стоят простые колонии, состоящие из 4-16 одинаковых жгутиковых клеток (некоторые представители родов Gonium и Pandorina), а завершают ряд виды рода Volvox, насчитывающие до 40 000 клеток. При этом появляется дифференциация клеток вдоль переднезадней оси колонии: клетки, расположенные впереди, имеют более развитые жгутики и глазки. Специализируются половые клетки, дающие мужские и женские гаметы, и особые клетки, обеспечивающие бесполое размножение.
Во всех рассмотренных выше случаях, за исключением ряда бурых водорослей, речь идет об организмах клеточного уровня организации, хотя многие из них имеют сложную дифференциацию ядер, того или иного рода многоядерное или даже поликлеточное состояние. В последнем случае они состоят из более-менее одинаковых вегетативных клеток, которые не дифференцируются на ткани (Серавин, 1991, 2000). Тканевая же организация, характерная для высших растений и животных, подразумевает решение стоящих перед организмом задач не за счет дифференцировок каких-либо внутриклеточных структур (например, формирования дополнительных органелл), а благодаря особой специализации целых групп клеток - тканей, каждая из которых выполняет определенную функцию. Организмы с тканевой организацией Л.Н.Серавин (1991) предлагает называть гетероклеточными, а не многоклеточными, чтобы отличать их от состоящих из нескольких клеток дотканевых эукариот.
Необходимо отметить, что становление тканевой организации у высших растений и животных проходило независимо. В результате у первых сформировались модульные живые системы, когда организм состоит из одной или нескольких разновидностей основных единиц, которые в определенном порядке сочетаются друг с другом (Марфенин, 1999). В случае растений отдельные модули представляют собой корни, целые стебли, ветви и листья. Причем потеря какого-либо модуля или даже многих модулей, как правило, не ведет к гибели целого организма. Наоборот, это в порядке вещей. Достаточно вспомнить хотя бы осенний листопад, сломанные ветки деревьев и т.п. Другое дело многоклеточные животные, особенно высшие их представители. Это унитарные системы с высоким уровнем интеграции, обеспечиваемым нервной системой и механизмами гуморальной регуляции. Модульная организация также встречается у животных, но только применительно к колониальным формам, таким, как колонии гидроидных и коралловых полипов, сифонофор и др.
Итак, приведенный анализ позволяет выделить внутри эукариот два крупных уровневых подразделения: дотканевые организмы клеточного уровня организации и тканевые, тканевого, соответственно, уровня организации, модульной, или унитарной природы. Тканевые, естественно, подразделяются на растения (начиная с мохообразных) и животных (начиная с кишечнополостных). К последним приближаются и губки. Однако именно приближаются. Дело в том, что организация губок не может быть названа тканевой в том смысле, который мы вкладываем в это понятие для прочих животных. Тканевые системы губок не устоялись. Их составляют весьма специализированные клетки, которые, однако, обладают значительной самостоятельностью, могут переходить от одного типа диффе-ренцировки к другому и менять при этом свою функциональную нагрузку, совершать перемещения внутри тела губки и т.п., то есть вести себя так, как никогда не ведет себя клетка, входящая в состав настоящих тканей. Губки - это что-то промежуточное между колониальной и тканевой организаций, но все же ближе к животным, чем к растениям, и традиционно считаются зоологическими объектами.
Дотканевые организмы представляют собой весьма пеструю смесь из простейших (в соответствии с зоологической номенклатурой) и водорослей (в соответствии с ботанической номенклатурой). Для них предлагается общее название протисты (протоксисты) (Whittaker, 1969; Маргелис, 1983). В эту группу включаются и так называемые грибоподобные протисты (протоктисты) - миксомицеты, оомицеты и другие зооспоровые грибы (Мюллер, Лёффлер, 1995). В контексте этих представлений не вполне решенным остается вопрос с бурыми водорослями, высокоспециализированные представители которых, как мы видели, по сути дела, следует считать тканевыми организмами. То же относится и к грибам. Они, в целом, стоят особняком и обладают целым рядом специфических особенностей (мицеллярная организация, хитиновая клеточная стенка и др.), которые достаточно отчетливо позволяют отделить их от протистов (см. ниже).
Подобная система взглядов привела к появлению пяти-царственной метасистемы живых организмов, которая впервые была сформулирована Уиттакером (Whittaker, 1969) (рис. 7) и получила широкое распространение (Маргелис, 1983; Карпов, 1990; Кусакин, 1995; Кусакин, Дроздов, 1998). Все прокариоты (архебактерии и эубактерии) объединяются в рамках царства Мопега, четко обособленного от всех эукариот. Среди последних различают царства Animalia (Животные), Plantae (Растения), Fungi (Грибы) и Protista (Протисты). К первому царству относятся организмы тканевой унитарной организации с гетеротрофным, преимущественно голозойным (захват твердых пищевых частиц внутрь тела) питанием. Царство Растений объединяет тканевые организмы модульной организации, преимущественно наземные, ав-тотрофные по способу питания, обладающие хлоропластами с хлорофиллом а и b и клеточной стенкой, в состав которой входит целлюлоза. Грибы - это мицеллярные гетеротрофы с сапрофитным способом питания и обладающие хитином в составе клеточной стенки. С растениями их сближает наличие клеточной стенки, способность к абсорбтивному извлечению необходимых веществ из растворов и чаще всего неподвижный наземный образ жизни вегетативных стадий. В то же время с животными грибы объединяют такие признаки, как гетеротрофия, присутствие хитина в клеточной стенке и мочевины в обмене веществ (Мираб-дуллаев, 1994). Три перечисленных выше царства эукариот отчетливо различаются и по своей экологической роли в биосфере: растения - это продуценты, животные - консументы, а грибы - редуценты.
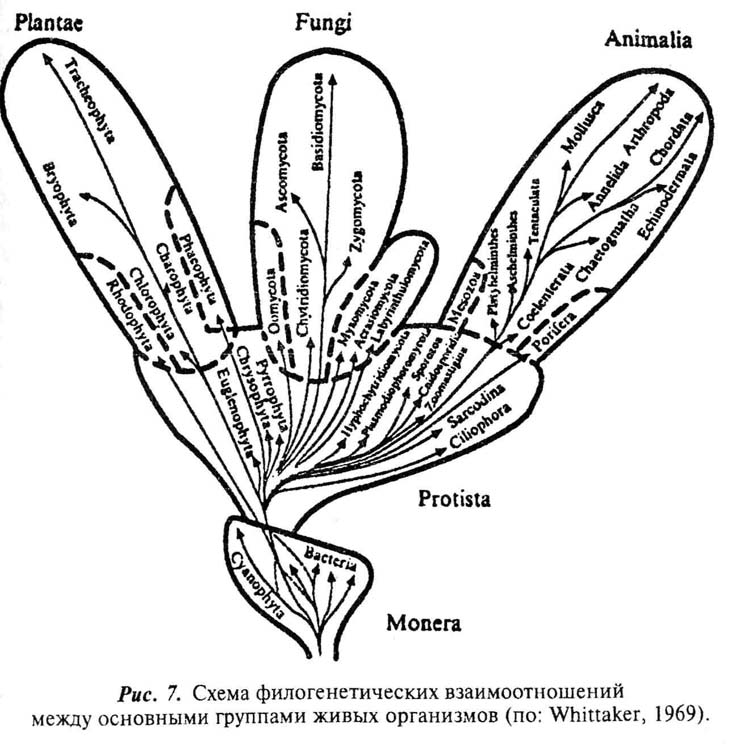
Всякая четкость пропадает, когда мы переходим к характеристике царства Протистов. Единственный более-менее надежный критерий - это дотканевой уровень организации. Причем в самых разнообразных вариантах, как то одноклеточное состояние, плазмодиальное, колониальное и т.п. (см. выше). Способы питания тоже самые различные. Это и авто-трофия, и разные модификации гетеротрофии и, наконец, миксотрофия, то есть способность одного и того же организма переходить от автотрофии к гетеротрофии и наоборот, в зависимости от параметров внешней среды. Практически все про-тисты подвижны хотя бы на одной из стадий жизненного цикла (у водорослей - это зооспоры и гаметы). Причем имеют место весьма разнообразные способы движения: при помощи жгутиков и ресничек, амебоидное и скользящее (диатомовые) движения. Для протистов характерно огромное разнообразие клеточных органелл, причем многие из них (экструсомы, сократительные вакуоли, гидрогеносомы, кинетопласты и др.), как уже отмечалось выше, не встречаются у высших эукариот. Кристы в митохондриях протистов, помимо характерной для животных, растений и грибов пластинчатой формы, могут быть трубчатые, пузырьковидные или ампуловидные (Полянский и др., 2000). У протистов мы сталкиваемся с многообразием вариантов строения хлоропластов (см. ниже), клеточных покровов, цитоскелетных структур (аксостили и т.п.) и др. Здесь выявляются все известные способы прохождения митоза и мейоза, самые разнообразные типы жизненных циклов, в которых причудливым образом сочетаются разные поколения, гаплоидные, диплоидные и полиплоидные стадии. Этот перечень можно продолжать еще очень долго.
Подобное многообразие протистов, учитывая все сказанное выше о путях становления жизни на Земле, не удивительно. Именно на уровне дотканевых эукариот, как мы видели, шла эволюция собственно эукариотной организации, приобретались разные клеточные системы, которые закреплялись затем у высших эукариот, использовались все возможности одноклеточного организма и апробировались разные варианты выхода за пределы этого состояния, шло становление полового процесса и испытывались самые различные типы жизненных циклов. Царство Протистов, таким образом, объединяет все результаты "эволюционных экспериментов" дотканевого уровня организации эукариот и потому не может быть цельным. В его рамках сложным образом переплетаются разные филогенетические линии, в том числе и те, которые в конечном итоге привели к появлению высших растений, животных и грибов. Поэтому искусственность царства Протистов в филогенетическом отношении никогда особых сомнений не вызывала. Оно сохранялось, да и сохраняется рядом систематиков до сих пор, поскольку наметить основные филогенетические линии и выделить крупные таксономические единицы, объединяющиеся на основе общности происхождения, вплоть до самого последнего времени оставалось задачей весьма нелегкой. Отсюда и многообразие трактовок одних и тех же данных, те кардинальные изменения в уже казалось бы сформировавшиеся представления, которые вносятся при появлении новых материалов, и, соответственно, то разнообразие мегасистем, которое можно обнаружить в литературе последних 10-20 лет.
Глава 7
ПОПЫТКИ ПОСТРОЕНИЯ "ФИЛОГЕНЕТИЧЕСКОЙ" МЕГАСИСТЕМЫ
Сразу следует оговориться, что рекомендовать какую-либо из так называемых филогенетических многоцарственных систем (число царств в некоторых из них достигает 18) пока не представляется возможным. На это, кстати, указывают и сами их создатели (Кусакин, 1995; Карпов, 2000). Несомненно, что эти системы в ближайшие годы будут подвергаться существенным переработкам. В то же время свет в конце туннеля уже виднеется. Объем накопленных к настоящему времени данных позволяет плодотворно использовать филогенетический подход и перейти в обосновании крупных таксономических единиц от гаданий на кофейной гуще к аргументированной дискуссии. Уже сейчас выделяется несколько достаточно четко обособленных групп, связанных общностью происхождения. Таксономический статус (царство, подцарство, тип или класс) этих групп может меняться от системы к системе, но их цельность и естественный характер выделения сомнений уже практически не вызывает. Это сигнализирует о том, что процесс создания метасистемы эукариот подходит к своему логическому завершению и уже в скором времени можно ожидать появления такой классификации, которая реально отражала бы ход эволюции эукариот и филогенетические взаимоотношения между разными их группами.
Ниже мы постараемся кратко охарактеризовать основные группы эукариотных организмов, выделение которых сейчас более-менее общепризнанно и проследить их филогенетические связи. Начнем с фотосинтезирующих эукариот, представления о филогении которых были заложены еще в трудах Па-шера (Pascher, 1918, 1931 - цит. по Старобогатов, 1986). Ближе к основанию филогенетического древа находятся красные водоросли, или багрянки, затем ствол фотосинтезирующих эукариот разделяется на две ветви. Одна - это зеленые водоросли и высшие растения, другая - золотистые, желто-зеленые, диатомовые и бурые. Примерно в район развилки попадают эвгленовые, криптофитовые и динофитовые. Подобное подразделение, а речь фактически идет о выделении трех основных группировок - красные, зеленые и хромофитовые - совпало с основными вариантами распределения хлорофиллов в хлоропластах. У красных это только хлорофилл а, у зеленых - а+b и а+с - у хромофитовых. Современные молекулярно-генетические и ультраструктурные данные подтвердили не случайный характер подобного распределения пигментов между разными группами водорослей и позволили пролить свет на происхождение хлоропластов в ходе эволюции эукариот.
Речь идет о симбиогенетическом происхождении, на что мы уже указывали в главе 3. Помимо особенностей структуры хлоропластов в пользу подобного предположения говорит и то обстоятельство, что хлорофилл а, характерный для хлоропластов всех растений, точно соответствует хлорофиллу а циано-бактерий и принципиально отличается от бактериохлорофилла прочих прокариот. Представить, что в ходе эволюции прокариот и эукариот столь сложное химическое соединение, как хлорофилл, возникло независимо, невозможно. Остается признать филогенетическую связь цианобактерий и хлоропластов. Известно, что цианобактерий легко входят в эндосимбиоти-ческие отношения с целым рядом протистов и берут на себя (не без пользы для своего хозяина) функцию фотосинтеза (Ха-усман, 1988; Карпов, 1990). Эндосимбиотические отношения могут заходить достаточно далеко. Так, у глаукофитовых, которые не обладают собственными хлоропластами, присутствующие в цитоплазме Эндосимбиотические цианеллы лишаются (частично или полностью) клеточной стенки и модифицируются до такой степени, что определить их систематическое положение среди цианобактерий оказывается невозможным (Седова, 1977; Карпов, 1990). Известны многочисленные примеры, когда в Эндосимбиотические отношения с протистами и многоклеточными животными (например, коралловыми полипами) вступают одноклеточные эукариотные водоросли (чаще всего динофитовые). Все это следует принять во внимание при рассмотрении структуры хлоропластов фотосинтезирующих эукариот, возможные пути эволюции которых были намечены Доджем (Dodge, 1979) и развиты в серии последующих исследований, среди которых выделяются многочисленные работы Ка-валье-Смита 80-90-х гг. (для обзора см.: Cavalir-Smith, 1999). Практически не вызывает споров эндосимбиотическое происхождение хлоропластов багрянок (рис. 8). В хлороплас-тах этих водорослей имеются одиночные, не контактирующие друг с другом тилакоиды (уплощенные цистерны, на которых локализованы воспринимающие свет пигменты). На их поверхности обнаруживаются мелкие тельца - фикобилисомы, в которых сосредоточены белки фикобилины. Они представляют собой дополнительные пигменты светособирающего комплекса и характерны для цианобактерий. От окружающей цитоплазмы хлоропласты красных водорослей отграничены двойной мембраной. Если предположить, что наружная из них ведет свое происхождение от мембраны фагоцитарной вакуоли, в которую когда-то был заключен эндосимбионт, то в остальном структура хлоропласта багрянок совпадает со строением цианобактерий.
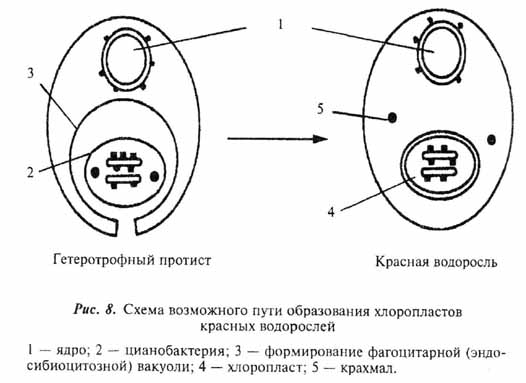
Мало сомнений вызывает и следующий шаг, который привел к появлению хлоропластов криптофитовых (рис. 9). Их окружают 4 мембраны, притом между второй и третьей имеется весьма обширное пространство, в котором обнаруживаются рибосомы, запасные питательные вещества и весьма своеобразная органелла - нуклеоморф. Название говорит само за себя. По своей структуре нуклеоморф напоминает ядро. Она окружена двойной мембраной с порами, имеет ДНК, небольшой участок РНК и делится при делении хлоропласта. Ее можно рассматривать как остаток ядра одноклеточной красной водоросли, которая когда-то вступила в эндосимбиоз с предком криптофитовых. Тогда в пространстве между второй и третьей мембраной локализуется то, что осталось от цитоплазмы этой эндосимбиотической багрянки. На родство с красными водорослями указывает и присутствие в попарно упакованных тилакоидах криптофитовых фикобилиновых белков, хотя они и не образуют здесь фикобилисом.
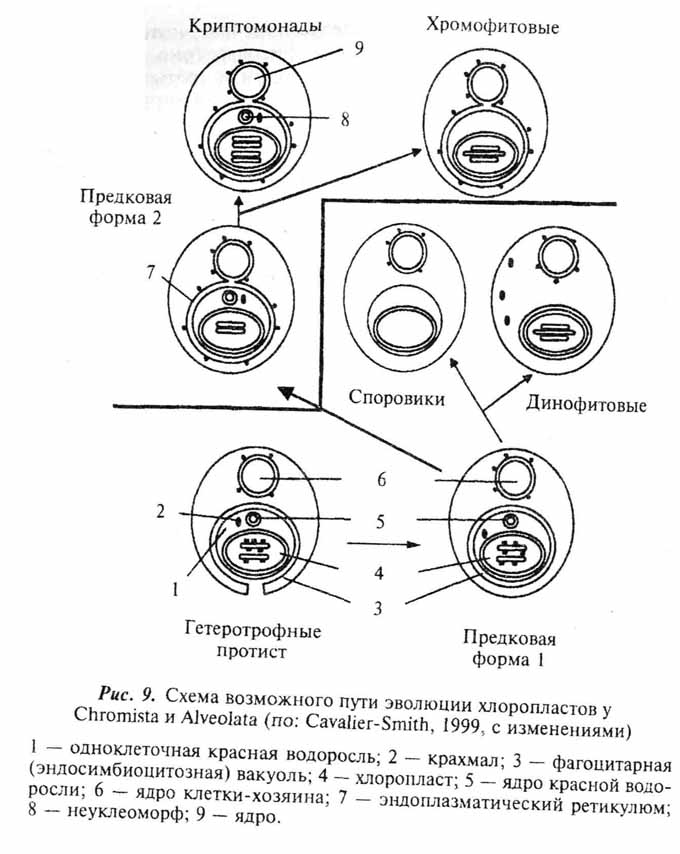
Механизм вторичного эндосимбиоза, то есть эндосимбиоза эукариотной одноклеточной водоросли, уже получившей хлоропласты в результате предшествующего эндосимбиоза с цианобактерией, с не имеющим еще хлоропластов эукариот-ным организмом, по-видимому, неоднократно задействовался в ходе эволюции. Такой путь формирования хлоропластов предполагается для всей обширной группы хромофитовых (рис. 9). У всех ее представителей хлоропласты окружают 4 мембраны, причем наружная представляет собой мембрану шероховатого эндоплазматического ретикулюма (ШЭПР), внутри которого оказываются хлоропласты. Потеря мембраны фагоцитарной вакуоли, некогда окружавшей эндосимбионта, и включение в цистерны ШЭПР можно рассматривать как адаптацию, облегчающую транспорт необходимых хлоропласту белков. Дело в том, что, как и в ходе эволюции митохондрий, кодирование значительной части необходимых для нормального функционирование хлоропласта белков постепенно перешло к ядерному аппарату. Синтез этих белков, соответственно, осуществляется на рибосомах ШЭПР. Их доставка непосредственно по его цистернам, минуя комплекс Гольджи, вполне может рассматривать как весьма полезное приспособление. Стоит отметить, что и наружная (четвертая) мембрана хлоропластов криптофитовых, обладающих нуклеоморфом, также предстаааяет собой мембрану ШЭПР.
Становление хлоропластов динофитовых тоже, по всей видимости, происходило в результате вторичного эндосимбиоза. У човременных представителей этой группы хлоропласты окружены тремя мембранами (рис. 9). Потерянной по предположению Кавалье-Смита (Cavalier-Smith, 1999), оказывается наружная мембрана вступившей в эндосимбиоз водоросли. Наружная же мембрана хлоропласта современных динофитовых берет свое начало от мембраны фагоцитарной вакуоли, окружавшей некогда эту эндосимбиотическую водоросль. Кавалье-Смит (Cavalier-Smith, 1999) связывает переход именно к такой структуре хлоропластов с особенностями транспорта обеспечивающих его работу белков из цитоплазмы клетки.
Рассмотренные выше группы водорослей, за исключением багрянок, обладают хлорофиллами а+с. Молекулы всех известных модификаций последнего (с1, с2, с3), в отличие от хлорофиллов а и b, не имеют фитолового конца и поэтому иногда именуются хлорофиллидами (см. Карпов, 1990). Предположение об их происхождении от настоящих хлорофиллов особых сомнений не вызывает. В то же время отсутствие у цианобактерий хлорофилла b долгое время было камнем преткновения для эндосимбиотической гипотезы происхождения хлоропластов зеленых растений. Однако в середине 70-х был выделен штамм цианобактерий, получивший название Prochloron, который обладал сочетанием хлорофиллов а и Ь. В дальнейшем было обнаружено еще несколько близкородственных прохлорону цианобактерий с хлорофиллами а+b. Интересен и тот факт, что если у цианобактерий тилакоиды в бактериоплазме лежат обычно отдельно друг от друга, то у содержащих хлоро-филлы а+b они формируют стопки, напоминающие в определенной степени граны в хлоропластах зеленых растений. Таким образом, происхождение хлоропластов зеленых водорос лей и высших растений в результате эндосимбиоза предковой формы с прохлорон-подобными цианобактериями кажется вполне правдоподобным. Эта точка зрения получила недавно еще одно молекулярно-генетическое подтверждение (Gogarten, 1995). В генах, кодирующих белки светособирающего комплекса у цианобактерий с хлорофиллами а+b, обнаружена делеция (потеря части генетического материала), и точно такая же делеция выявлена в хлоропластах зеленых водорослей и высших растений. В то же время она отсутствует в генетическом материале хлоропластов у исследованных групп хромофитовых и красных водорослей.
Следует вспомнить еще об одной группе фотосинтезирую-щих эукариот с хлорофиллами а+b. Это эвгленовые, хлоропласты которых окружены тремя мембранами. Их происхождение связывают с механизмом вторичного эндосимбиоза, сходного с описанным выше для динофитовых (Cavalier-Smith, 1999) (рис. 10). Однако, в отличие от последних, роль эндосимбион-та играли какие-то одноклеточные зеленые, а не красные водоросли. Остается упомянуть весьма своеобразные зеленые водоросли Chlorarachnlon, которых Кавалье-Смит (Cavalier-Smith, 1999) склонен сближать с эвгленовыми. Хлоропласт этих водорослей окружен 4 мембранами и, как у криптофито-вых, между второй и третьей мембраной помещается нуклео-морф, что служит достаточно веским указанием на происхождение этой органеллы в результате вторичного эндосимбиоза.
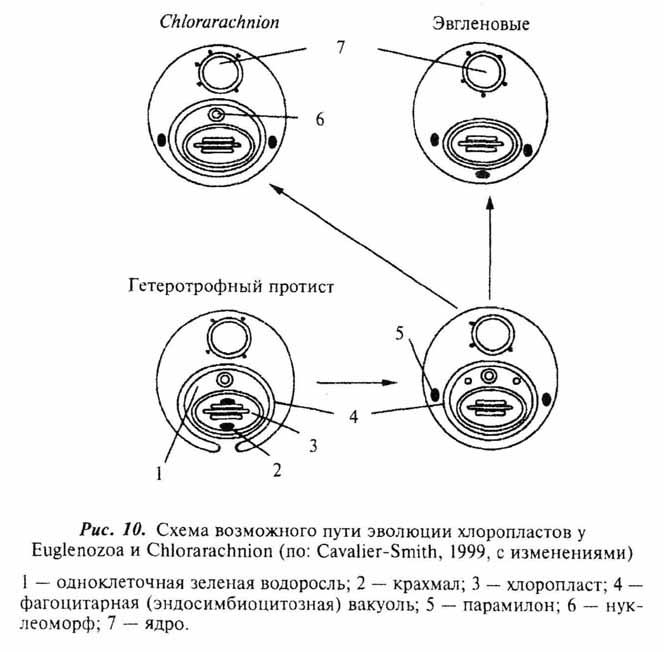
Подводя итог приведенному выше анализу структуры и возможных путей происхождения хлоропластов в разных группах фотосинтезирующих эукариот, можно сделать однозначный вывод: хлоропласты возникали независимо и неоднократно. Это служит еще одним доводом в пользу того, что сам факт наличия хлоропластов нельзя рассматривать как признак, говорящий о филогенетической общности. Поэтому выделение царства Растений в том понимании, которое вкладывали в него еще со времен Аристотеля, то есть как объединение всех фотосинтезирующих эукариот, с позиций филогенетического подхода представляется абсолютно неправомочным. В то же время среди растительных организмов выделяется несколько естественных групп, связанных филогенетическим родством (рис. 11).
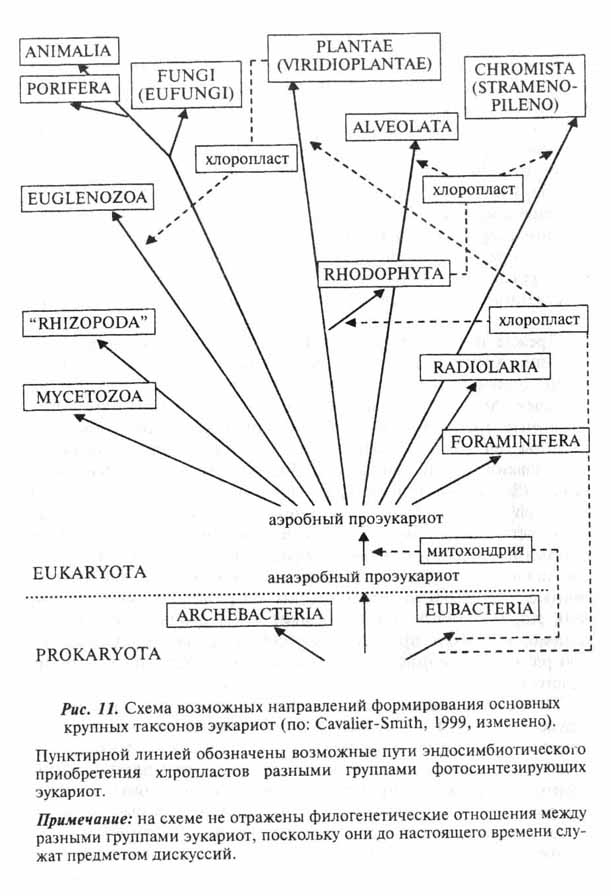
Прежде всего это зеленые водоросли, харовые и высшие растения. Хлоропласты всех этих организмов устроены совершенно одинаково и их общее происхождение сомнений не вызывает. Можно только согласиться с объединением перечисленных групп в рамках более крупной таксономической единицы (возможно, в ранге царства), для которой предлагается название Viridioplantae, или Plantae (растения). Нередко в состав Plantae включают красные водоросли, или багрянки (Rhodophyta). Подобное объединение весьма спорно. Достаточно вспомнить описанное выше строение хлоропластов, в которых присутствует только хлорофилл а, особенности строения талломов, характер протекания митоза и другие специфичные особенности организации багрянок (Саут, Уиттик, 1990; Карпов, 1990). Все это ставит их несколько особняком. Видимо, багрянки представляют собой самостоятельную линию развития эукариот, достаточно рано отделившуюся от их общего ствола.
Не менее естественным, чем зеленые растения, выглядит и выделение группы хромофитиовых или хромистов (Chromista), в рамках которой объединяются все водоросли с хлорофилла-ми а+с. По этому признаку к хромистам причисляют и крип-тофитовые, однако, многие исследователи предпочитают рассматривать их в качестве самостоятельной группы высокого ранга. Следует отметить, что по особенностям строения жгутикового аппарата, митохондрий и молекулярно-генетическим данным к хромистам относят и некоторые группы бесцветных организмов. В первую очередь, это зооспоровые грибы (оомице-ты и другие бывшие зооспоровые грибы, кроме хитридиевых), уже упоминавшиеся выше опалины и ряд других более мелких таксонов (Карпов, 2000). Поскольку в таком составе группа оказывается представленной не только фотосинтезирующими, но и гетеротрофными организмами, то ее в последнее время предлагают переименовать в страминопилов (Stramenopiles), чтобы избежать в названии акцента на наличие хлоропластов. Это название, надо отметить, пока еще не получило широкого хождения.
Вполне устоявшейся группой высокого таксономического уровня следует признать и альвеолят (Alveolata). В состав этого таксона входят такие еще недавно казавшиеся далеко отстоящими друг от друга группы протистов, как инфузории (Ciliophora), споровики (Sprozoa или Apicompiexa) (паразитические простейшие, в числе которых малярийный плазмодий, кокцидии и др.) и динофитовые (Dinophyta). Первые два таксона представлены исключительно гетеротрофами, динофитовые преимущественно автотрофны, но значительное число их видов переходит к хищничеству, детритофагии или паразитизму, что влечет за собой потерю хлоропластов. В то же время у споровиков выявлены пластиды (хлоропласты, лишенные ти-лакоидов и, соответственно, белков светособирающего комплекса, служащих для запасания питательных веществ), которые одеты тремя мембранами. Их происхождение может быть связано со вторичным эндосимбиозом с одноклеточной красной водорослью какого-то гетеротрофного протиста, который явился общим предком для споровиков и динофитовых (Cavalier-Smith, 1999). Свое название Alveolata получили из-за особенностей организации их покровов. Под наружной плазматической мембраной у них локализуется слой уплощенных пузырьков - альвеол (Карпов, 1990).
Другая группа, включающая в себя также разнородных по способу питания протистов, носит название Euglenozoa. В ее состав входят уже не раз упоминавшиеся эвгленовые, среди которых есть и автотрофы, и гетероторфы, и миксотрофы. Кроме них, к Euglenozoa относится и почти целиком паразитическая группа жгутиконосцев - кинетопластид (Kinetoplastida), получивших свое название от наличия прокрашивающейся ядерными красителями структуры - кинетопласта. Современные исследования показали, что она представляет собой участок гигантской митохондрии, содержащий множество кольцевых молекул ДНК (Заварзин и др., 1992). К кинетопластидам принадлежат в том числе возбудители (трипанозомы, лейшма-нии) ряда опасных заболеваний человека, таких как сонная болезнь, кала-азар, пендинская язва и др.
Все остальные выделяемые в настоящее время крупные таксономические группы эукариот представлены только гете-ротрофами. Среди них, конечно, прежде всего следует назвать Metazoa (многоклеточные, а вернее, разноклеточные - см. главу 6), которые включают всех тканевых животных, начиная с кишечнополостых, и губок (Porifera). В этом случае границы таксона практически совпадают с намеченными при использовании уровневого подхода (см. главу 5). В последние годы к Metazoa стали относить и обширную группу паразитических организмов - миксоспоридий, которые всегда рассматривались зоологами как представители типа Protozoa (простейшие). Вегетативное тело этих паразитов представлено многоядерными плазмодиями, в которых формируются многоклеточные споры. Они обладают специализированным аппаратом, который выстреливает зародыш прямо внутрь тканей хозяина. Тщательное изучение этого аппарата показало, что он сходен с аналогичной структурой стрекательных капсул кишечнополостных. Кроме того, детальное исследование паразитических полипов Polypodium (на паразитической стадии образуют плазмодиоподобную структуру в икре осетровых рыб) прояснило возможный путь перехода от организации типичного кишечнополостного к характерной для миксоспоридий.
С миксоспоридиями связана и еще одна недавняя зоологическая сенсация. Всегда считалось, что они паразитируют на вегетативной стадии в тканях преимущественно костистых рыб, а споры рассеиваются в воде и захватываются вместе с ней рыбами при дыхании и питании. В то же время представители близкородственной группы простейших - актиномикси-дии паразитируют преимущественно в бентосных кольчатых червях (стадия плазмодия) и так же рассеивают во внешнюю среду многоклеточные споры, имеющие, однако, иное строение, чем у миксоспоридий. Недавно выполненные тонкие эксперименты показали, что миксоспоридий и актиномиксидии не два самостоятельных таксона, а их представители суть ни что иное, как разные части одних и тех же жизненных циклов, причем обе эти части обладают известной самостоятельностью. Споры миксоспоридий, попадая в кольчецов, могут давать начало плазмодию, продуцирующему споры актиномик-сидного типа. Последние способны заражать рыб, формировать плазмодий, внутри которого развиваются типичные споры миксоспоридийного типа. В то же время обе части жизненного цикла (споры миксоспоридийного типа - рыбы - споры миксоспоридийного типа и споры актиномиксидного типа - кольчецы - споры актиномиксидного типа) могут многократно реализовываться совершенно независимо друг от друга. Следует добавить, что в обеих этих частях имеет место половой процесс и смена гапло- и диплофаз. Таким образом, жизненный цикл миксоспоридий в полном объеме претендует на место самого сложного среди современных эукариот.
Еще относительно недавно происхождение многоклеточных связывали с колониальными зелеными жгутиконосцами (Иванов, 1973). Однако это не подтвердилось современными молекулярно-генетическими данными, которые достаточно отчетливо указывают на родство животных с грибами. В состав последних, как мы уже писали выше, включаются только аско-мицеты, зигомицеты, базидиомицеты и хитридиевый, и во избежание путаницы грибы в таком составе часто обозначают как Euiungi, а не Fungi. Хитридиевые представляют собой небольшую группу микроскопических грибов, в ходе бесполого размножения которых образуются одножгутиковые зооспоры. Ультраструктурные исследования этих зооспор показали, что их жгутиковый аппарат устроен сходно с таковым хоаноцитов - клеток, выстилающих жгутиковые камеры губок, и совсем не так, как у зеленых жгутиконосцев (Карпов, Ефремова, 1994). Таким образом, хитридиевые (или родственные им вымершие формы) претендуют на роль предков для Metazoa, а грибы, соответственно, оказываются гораздо ближе к животным, чем к растениям, хотя традиционно они до сих пор считаются ботаническими объектами.
В состав Eufungi стали включать и микроспоридий (Microsporidia). На вегетативной стадии они образуют небольшие плазмодии или мицелиеподобные структуры внутри клеток животных и растений. Распространение паразитов происходит при помощи спор, обладающих сложным аппаратом внедрения, благодаря которому спороплазма выстреливается внутрь клетки-хозяина. Аппарат этот устроен по иному принципу, чем у рассматривавшихся выше миксоспоридий и акти-номиксидий и не имеет ничего общего со стрекательным аппаратом кишечнополостных. Особый интерес к микроспоридиям вызывало то обстоятельство, что у них отсутствуют митохондрии. До недавнего времени считалось, что это первичный признак и что линия микроспоридий обособилась от остальных эукариот до приобретения ими митохондрий. Поэтому их нередко выделяли в самостоятельный таксон крупного ранга вплоть до царства. Однако подобные взгляды не подтвердились (Muller, 1997; Hasegawa, Hashimoto, 1999). В ядерном геноме микроспоридий были обнаружены последовательности, кодирующие некоторые митохондриальные белки. Это однозначно свидетельствует, что предок микроспоридий обладал митохондриями, а их вторичная утрата связана, очевидно, с переходом к внутриклеточному паразитизму, то есть обитанию в анаэробных условиях. Молекулярно-генетические данные также подтверждают близость микроспоридий и грибов.
Кроме оомицетов и некоторых более мелких групп так называемых зооспоровых грибов, которых в соответствии с современными представлениями относят к хромистам (страме-нопилам), к грибам раньше относили и миксомицетов. В настоящее время стало очевидным, что они обладают целым рядом ультраструктурных признаков, сближающих их с некоторыми пресноводными жгутиконосцами (церкомонады), вместе с которыми они и образуют компактную группу Mycetozoa. Следует отметить, что, в отличие от настоящих грибов, мице-тозои питаются голозойным способом, заглатывая различных бактерий и протестов.
Обширную и разнородную группу представляют собой так называемые корненожки - "Rhizopoda". Заключение этого названия в кавычки не случайно и объясняется тем обстоятельством, что группа эта чрезвычайно гетерогенна по своему составу. Она объединяет одноклеточных, или плазмодиальных, протистов, обладающих, как правило, амебоидной подвижностью. Жгутиковые стадии, если и имеются, то весьма кратковременны. У большинства же корненожек они отсутствуют на всех стадиях жизненного цикла. Амебоидная организация дает очень мало морфологических привязок даже на ультраструктурном уровне, на которых можно было бы обосновать более-менее естественную классификацию этих организмов. Поэтому корненожки до сих пор остаются сборной группой, включающей, очевидно, разнородных по своему происхождению амебоидных организмов. Что стало совершенно отчетливо на сегодняшний день - это выведение за ее пределы фораминифер (Foraminifera). Это хорошо очерченная и многочисленная группа протистов, обладающих раковиной разнообразного строения и конфигурации. Стенки раковин пронизывают поры, через которые наружу высовываются специализированные отростки цитоплазмы - ретикулоподии. В их центральной части проходит пучок микротрубочек, которые, вращаясь друг за другом наподобие храповика, обеспечивают движение поверхностных слоев цитоплазмы ретикулоподии. При помощи ретикулоподии фораминиферы перемещаются по субстрату и захватывают добычу.
Несомненно, особняком стоит и обширная сборная группа морских протистов - радиолярий (Radiolaria), обладающих внутренним минеральным скелетом, который часто формирует причудливые, геометрически правильные структуры. Радиолярии имеют, как правило, одно крупное полиплоидное (полигеномное) ядро, сложно дифференцированную цитоплазму, дающую многочисленные нитевидные выросты (аксоподии), отходящие во все стороны от тела. В аксоподиях, так же как и в ретикулоподиях фораминифер, имеются центральные пучки микротрубочек, образующих сложные пространственные структуры, которые принципиально отличаются от обнаруживаемых у фораминифер. Поверхностный слой цитоплазмы аксоподии находится в непрерывном движении и обеспечивает транспортировку мелких пищевых частиц к центральной цитоплазме радиолярии.
Заслуживает упоминания вопрос о выделении группы Archezoa, которую Кавалье-Смит сохраняет и в своих последних вариантах системы эукариот (Cavalier-Smith, 1999). В ее состав первоначально включались протисты (микроспоридии, дипломонады, трихомонады и некоторые другие), которые лишены митохондрий. О микроспоридиях речь уже шла. Дипломонады и трихомонады также ведут паразитический образ жизни и обладают своеобразно устроенным жгутиковым аппаратом, в состав которого входят 4 жгутика, 4 кинетосомы и ряд ассоциированных структур. К первым относятся печально известные лямблии, ко вторым не менее широко распространенный паразит человека Trichomonas vaginalis. Отсутствие митохондрий у архезоев рассматривалось как древний первичный признак и служило указанием на раннее, до приобретения митохондрий, их отделение от общего ствола эукариот. Подтверждали эту точку зрения и результаты анализа рибосомальных РНК, в соответствии с которыми дипломонады и трихомонады оказывались у основания филогенетического древа. Последующие исследования показали, что не только у микроспоридий (см. выше), но и у всех остальных архезоев в ядерном геноме существуют последовательности, кодирующие некоторые митохондриальные белки (Hasegawa, Hashimoto, 1999). Иными словами, отсутствие митохондрий у них следует считать вторичным признаком и связано с паразитическим образом жизни. Дополнительные молекулярные исследования, в которых были задействованы не только рибосомальные РНК, но и участки ядерной ДНК, кодирующие важнейшие группы белков (актины, тубулины и др.), показали, что группа Archezoa - сборная, и ее представители достаточно легко распределяются среди известных групп эукариот. Микроспоридии, как уже отмечалось, отошли к грибам, а трихомонады и дипломонады, естественно, вошли в состав группы Polymastigota и т.д. (Карпов, 2000). Таким образом, валидность выделения архезоев в качестве самостоятельной линии эволюции эукариот представляется сомнительной и не подтверждается новыми данными.
Приведенный выше краткий обзор, конечно, не охватывает всех известных эукариот. Многие, даже относительно многочисленные их группы, положение которых в системе еще во многом не понятно, остались за рамками изложения. Таксономический же статус охарактеризованных нами групп пока еще дискуссионен, хотя их реальность как естественных филогенетических группировок сомнений уже не вызывает. В своей последней системе Кавалье-Смит (Cavalier-Smith, 1999) предлагает разделить все живые существа на 6 царств: Bacteria (все прокариоты), Animalia (многоклеточные, точнее, гетерокле-точные, животные), Fungi (в смысле Eufungi), Plantae (в смысле Viridioplantae, плюс красные водоросли), Chromista (включая и альвеолят) и Protozoa (все, что не вошло в предыдущие пять царств). Если с первой пятеркой еще с очень большой натяжкой (красные водоросли все же представляют отдельную от зеленых линию развития, то же можно сказать и о альвеолятах по отношению к остальным хромистам) можно согласиться, то царство Protozoa, в постулируемом Кавалье-Смитом (Cavalier-Smith, 1999) составе, представляет собой такую смесь разных филогенетических линий, которая сравнима разве что с царством Protista, выделенном на основе уровневого подхода (см. главу 5). Тем не менее, мегасистема живых организмов уже приобретает некоторые черты реальности и, повторяя уже сказанное в начале главы, можно прогнозировать скорое появление достаточно приемлемой классификации.
СПИСОК ЛИТЕРАТУРЫ
Айдилл С.П. Анчоусовый кризис // Наука об океане. М.: Прогресс. 1981. С. 279-294.
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Роберте К., Уотсон Дж. Молекулярная биология клетки. Т. I-V. М.: Мир. 1986.
Видаль Г. Древнейшие эукариотические клетки // В мире науки.
1984, № 4. С. 14-24.
Виноградова К.Л. Отдел красные водоросли (Rhodophyta) // Жизнь растений. Т. 3. М.: Просвещение, 1977. С. 192-250.
Георгиев Т.П. Гены высших организмов и их экспрессия. М.: Наука. 1989. 254 с.
Грин П., Стаут У., Тейлор Д. Биология. Т. 3. М.: Мир. 1990, 374с.
Громов Б.В. Строение бактерий. Л.: Изд-во Ленингр. гос. ун-та.
1985. 200 с.
Дикерсон Р.Е. Химическая эволюция и происхождение жизни //
Эволюция. М.: Наука. 1981. С. 67-107.
Докинз Р. Эгоистичный ген. М.: Мир. 1993.
Заварзин А.А., Харазова А.Д., Молитвин М.Н. Биология клетки. СПб: изд-во СПбГУ. 1992.
Иванов А.В. Происхождение многоклеточных животных. Л.: Наука. 1973.
Инге-Вечтомов С.Г. Генетика с основами селекции. М.: Высшая школа. 1989.
Карпов С.А. Система протистов, Омск. 1990.
Карпов С.А. Система протистов и проблемы мегасистематики // Протесты. Часть 1. Спб.: Наука. 2000. С. 123-137.
Карпов С.А., Ефремова СМ. Ультратонкое строение жгутикового аппарата губки Ephydatia fluviatilis // Цитология. 1994. Т. 36, № 5.
Кернс-Смит А.Д. Первые организмы // В мире науки. 1985. № 8. С. 46-56.
Кимура М. Молекулярная эволюция: теория нейтральности. - М., Мир. 1985.
Крылов И.Н. Древнейшие следы жизни // Природа. 1985. № 9. С. 68-76.
Кусакин О. Г. Кризис в мегаттаксономии и пути его преодоления. 1. От традиционных полифилеткческих систем евкариот к многоцарственным монофилетическим // Биол. моря (Владивосток). 1995. Т. 21, № 4. С. 236-250.
Кусакин О.Г., Дроздов А.Л. Филема органического мира. 2. Прокариоты и низшие эвкариоты. СПб: Наука. 1998.
Лемеза Н.А., Камлюк Л.В., Лисов Н.Д. Биология в экзаменационных вопросах и ответах. М.: Рольф, Айрис-пресс. 1999.
Марфенин Н.Н. Концепция модульной организации в развитии / / Журнал общей биологии. 1999. Т. 60, №1. С.6-17.
Маргелис Л. Роль симбиоза в эволюции клетки. - М., Мир. 1983.
Медников Б.М. Новая макросистематика // Биология в школе. 1997. № 5. С. 5-9.
Михалевич В.И. Тип Foraminifera // Протисты. Часть 1. Спб.: Наука. 2000. С. 533-623.
Мирабдуллаев И.М. О происхождении и положении грибов в системе органического мира // Успехи соврем, биологии. 1994. Т. 114, вып. 1. С. 30-41.
Мусил Я., Новикова О., Кунц К. Современная биохимия в схемах. М.: Мир. 1984.
Мюллер Э., Лё'ффлер В. Микология. М.: Мир. 1995.
Нолл Э.Х. Конец протерозойского зона // В мире науки. 1991. № 12. С. 26-34.
Опарин А. И. Возникновение и начальное развитие жизни. М.: Медицина. 1966.
Полянский Ю.И. О своеобразных чертах прогрессивной эволюции на клеточном уровне организации // Журнал общей биологии. 1971. Т. 32, № 5. С. 541-548.
Полянский Ю.И., Суханова К.М., Карпов С.А. Общая характеристика протистов // Протисты. Часть 1. Спб.: Наука. 2000. С. 145-190.
Райков И.Б. Ядро простейших. Л.: Наука. 1978. Розанов А.Ю., Федонкин М.А. Проблема первичного биотопа эв-кариот // Экосистемные перестройки и эволюция биосферы. Выпуск 1. М.: Недра. 1994. С. 25-32.
Саут Р., Уиттик А. Основы альгологии. М.: Мир. 1990.
Седова Т.В. Сожительство водорослей с другими организмами // Жизнь растений. Т. 3. М.: Просвещение. 1977. С.12-11.
Серавин Л.Н. Вариации в строении аксонемы жгутика и происхождение этой органеллы // Цитология. 1985. Т. 27. С. 971-985.
Серавин Л.Н. Происхождение эукариотической клетки. I. Исторические истоки и современное состояние концепций симбиотичес-кого и аутогенного происхождения клетки // Цитология. 1986а. Т. 28, № 6. С.563-574.
Серавин Л.Н. Происхождение эукариотической клетки. П. Критический анализ симбиотической (экзогенной) концепции // Цитология. 19866. Т. 28, № 7. С. 659-669.
Серавин Л.Н. Происхождение эукариотической клетки. III. Некоторые принципы морфофункциональной организации эукариот-ной клетки // Цитология. 1986в. Т. 28, № 8. С. 779-789.
Серавин Л.Н. Происхождение эукариотической клетки. IV. Общая гипотеза аутогенетического происхождения эукариот // Цитология. 1986г. Т. 28, № 9. С. 899-910.
Серавин Л.Н. Существует ли в действительности такой таксон - подцарство Protozoa? // Биол. науки. 1989. Т. 7. С. 4-3.
Серавин Л.Н. Основные этапы развития клеточной теории и место клетки среди живых систем // Цитология. 1991. Т. 33, № 12. С. 3-27.
Серавин Л.Н. Основной тип питания первичных организмов (протобионтов) и его дальнейшая эволюция // Тр. Спб, общ-ва естественен. 1994. Т. 90, вып. 1. С. 88-96.
Серавин Л.Н. Современные представления о месте губок в системе эукариот // Вестник СПбГУ. Сер. 3. 1997, вып. 3 (№ 17). С. 34-41.
Серавин Л.Н. Пути эволюции протистов // Протисты. Часть 1. Спб.: Наука. 2000. С. 138-145.
Соколов Б.С. Вендский период в истории Земли // Природа. 1984. № 12. С. 3-18.
Спирин А.С. Биосинтез белков, мир РНК и происхождение жизни // Вестник Российской Академии Наук. 2001. Т. 71, № 4. С. 320- 328.
Старобогатов Я. И. К вопросу о числе царств эукариотных организмов // Труды Зоол. ин-та АН СССР. 1986. Т. 144. С. 4-25.
Татаринов Л.П. Очерки по теории эволюции. - М., Наука.
1987.
Тыщенко В.П. Введение в теорию эволюции. - СПб., изд-во
СПбГУ. 1992.
Фолсом К. Происхождение жизни. - М., Мир. 1982.
Федонкин М.А. Крупнейшее месторождение вендской фауны // Природа. 1981. № 5. С. 94-102.
Федонкин М.А. Бесскелетная фауна венда и ее место в эволюции Metazoa. - М.: Наука. 1987.
Федонкин М.А. Второе рождение кимбереллы // Природа. 1998. № 1. С. 3-10.
Федонкин М.А. Холодная заря животной жизни // Природа. 2000. № 9. С. 3-11.
Хаусман К. Протозоология. М.: Мир. 1988.
Хорган Д. У истоков жизни // В мире науки. 1991. № 4. С. 69-79.
Чайковский Ю.В. Макросистема не похожая на другие // Биология в школе. 1998. № 6. С. 19.
Чумаков Н.М. Климатический парадокс позднего докембрия // Природа. 1992. № 4. С. 34-41.
Шипунов А. Б. Сколько царств живых организмов? Биология в школе. 1997. № 4. С. 5-11.
Шопф Д.У. Эволюция первых клеток // Эволюция. М.: Наука. 1981. С. 109-147.
Bell М.А. Origin of metazoan phyla: Cambrian explosion or proterozoic slow burn? // TREE. 1997. V. 12, N 1. P. 1-2.
Cavalier-Smyth T. Eukaryote kingdoms: seven or nine? // BioSystems. 1981. V. 14. P. 461-484.
Cavalier-Smyth T. The origin of Eukaryote and Archebacterial cells. Endocytobiology. III. // Ann. N. Y. Ac. Sci. 1987. V. 503. P. 17-54.
Cavalier-Smyth T. Principles of protein and lipid targeting in secondary symbiogenesis: euglenoid, dinoflagellate and sporozoan plastid origins and the eukaryote family tree. J. Eukaryot. Microbiol. 1999. V. 46. N 4, p. 347-366.
Chang S. Prebiotic synthesis in planetary environments // In: The Chemistry of Life's Origins. J.M. Greenberg et al. (eds.). Kluwer Acad. Publ.: Dordrecht, The Netherlands. 1993. P. 259-299.
Chang S. Planetary environments and the origin of life // Biology Bulletin. 1999. V. 196. P. 308-310.
Chyba C.F. Buried beginning // Nature. 1998. V. 395. P. 329-330.
Chyba C.F., Sagan C. Endogenous production, exogenous delivery, and impact-shock synthesis of organic molecules: an inventory for the origins of life // Nature. 1992. V. 355. P. 125-131.
Corlis J.O. Protistian phytogeny and eukaryogenesis // Intern. Rev. Cytol. 1987. V. 100. P. 313-370.
De Duve C. The birth of complex cell // Scientific American. April 1996. P. 40-45.
Dodge J.D. The phytoflagellates: fine structure and phylogeny // Biochemistry and physiology of Protozoa. V.I. London: Academic Press. P. 7-57.
Gilbert W. The RNA world // Nature. 1986. V. 319. P. 618.
Gogarten J.P. The early evolution of cellular life // TREE. 1995. Vol. 10, N 4. P. 147-151.
Gray M.W., Burger G., Cedergren R., Golding G.B., Lemeieux C., Sankoff D., Turmel M., Lang B.F. A genomic approach to mitochondrial evolution // Biological Bulletin. 1999. Vol. 196. P. 400-402.
Haeckel E. Generelle Morphologic der Organismen. I. Allgemeine Anatomic der Organismen. II. Allgemeine Entwickingsgeschichte der Organismen. Berlin: G. Reimer. 1866.
Haeckel E. Das Protistenreich. Leipzig: Gunther. 1878.
Hasegawa M., Hashimoto T. Phylogenetic position of amitochondriate protists in the evolution of eukaryotes // Biological Bulletin. 1999. Vol. 196. P. 389-391.
Lang B.F., Seif E., Gray M.W., O'Kelly C.J., Burger G. A comparative genomics approach to the evolution of eukaryotes and their mitochondria // The Journal of Eukaryotic Microbiology. 1999. V.46, N 4. P.320-326.
Muller M. What are the Microsporidia? // Parasitology Today. 1997. V. 13, N 12. P. 455-456.
Orgel L.E. The origin of life - a review of facts and speculations // Trends in Biochemical Sciences. 1998. V. 23. P. 491-495.
Reysenbach A.-L., Seitzinger S., Kirshtein J., McLauglin E. Molecular constraints on a high-temperature evolution of early life // Biological Bulletin. 1999. Vol. 196. P. 367-372.
Schopf J. W. Deep divisions in the tree of life - what does the fossil record reveal? // Biological Bulletin. 1999. Vol. 196. P. 351-355.
Schidlowski M. Life in Isua times? A frontier revision // Origin of Life and Evolution of Biosphere. 1994. Vol. 24, N 2/4. P. 267-268.
Turner S., Pryer K.M., Miao V.P. W., Palmer J.D. Investigating deep phylogenetic relationships among Cyanobacteria and plastids by small subunit rRNA sequence analysis // The Journal of Eukaryotic Microbiology. 1999. V.46, N 4. P. 327-338.
Whittaker R.H. New concept of kingdoms of organisms // Science.
1969. V. 183. P. 150-159.
Wray G.A., Levinton J.S., Shapiro L.H. Molecular evidence for deep pre-Cambrian divergences among metazoan phyla // Science. 1996. N 274. P. 568-573.